|
INTEGRATED OVERVIEW OF FISHERIES OF THE BENGUELA CURRENT REGION A synthesis commissioned by the United Nations Development Programme (UNDP) as an information source for the Benguela Current Large Marine Ecosystem (BCLME) Programme Authors
EXECUTIVE SUMMARY This Report gives an overview of the major living resources of the Benguela Current Ecosystem, and of the research which has been conducted on them by the three countries bordering the system (Angola, Namibia and South Africa) and foreign partners, as input to management of these resources. It treats the resources on a regional basis, and highlights trans-boundary problems which need to be addressed through a regional rather than a national approach. The report amplifies and updates recent summaries of scientific information on the resources, and adds material on the legal framework and structures under which they are managed in each of the three countries. The distribution and habitats of the major species exploited by the purse seine, trawl, crustacean, linefish and artisanal fisheries of the region, and of the seal and main seabird populations, are briefly described and discussed with particular reference to questions of stock separation and the connection between apparently different populations. It is pointed out that a number of commercially-important species (e.g. hake, horse mackerel, deep-sea red crab, tuna and, probably to a lesser extent, sardine and anchovy) are distributed or move seasonally across national boundaries, requiring regional compatability in the research and management of these resources. The life histories of the major exploited species are briefly outlined, with emphasis on temporal and spatial spawning patterns, the dispersal of the early-life stages, migration patterns of recruits and adults, and diet (particularly as it relates to potential competition between species). The broad picture to emerge is that the important offshore resources in the southern Benguela spawn over various parts of the Agulhas Bank, and depend to a greater or lesser extent on the equatorward jet current between Cape Point and Cape Columbine to transport the early life stages to the West Coast, where much of the recruitment takes place. Pelagic recruits return to the Agulhas Bank in the poleward counter current close to the coast. Species which become more demersal with age (e.g. hake and horse mackerel) tend to move into deeper water as they move south in the early stages, resulting in complex movements between the West and South Coasts and more widespread spawning areas. In the northern Benguela, spawning of many of the important species is concentrated in northern Namibia/southern Angola, and is probably influenced by movements of the Angola-Benguela front. Juveniles spawned in this area tend to move south close to the coast and to return northwards farther offshore when older. The history and current status of the major fisheries of the region are discussed, concentrating on catch trends and changes in the nature of the fisheries. Over the past 30 to 40 years total catches in the south-east Atlantic have declined from a peak of more than 3 million tonnes in 1968 to levels of around 1 million tonnes per year in the 1990s. The most pronounced features in Namibia and South Africa have been the major decline in catches of South African and Namibian sardine in the mid- and late 1960s respectively (followed by a dramatic further decline in Namibia in the mid-1970s), the major decline in the West Coast rock lobster resource (particularly off Namibia) to levels well below those in 1960, and the major reduction in hake and horse mackerel catches off Namibia in the 1990s, due largely to the withdrawal of foreign fishing fleets after Independence in 1990. Off Angola, the most notable feature is the sharp reduction in industrial catches of all the most important species (e.g. sardinellas, horse mackerel and deep-water prawns), due largely to the major reduction in foreign fishing effort from 1985 onwards. Changes in the abundance and distribution of the major resources, including seals and seabirds, as revealed by acoustic, trawl and aerial surveys and catch-based analytical methods, are discussed. VPA estimates confirm the dramatic decline of Namibian sardine in the late 1960s and the subsequent further sharp decline in the mid-1970s. Acoustic surveys since 1990 show that the population dropped to its lowest ever level in 1995 (co-incident with a major Benguela Nińo in the northern Benguela), but that it has recovered somewhat since then. Acoustic surveys of sardine and anchovy in South African waters indicate that there has been a gradual increase in sardine abundance since the mid-1980s (although probably not to pre-collapse levels), and show two cycles in anchovy spawner biomass, with peaks in 1986 and 1991. The distribution of Namibian sardine has moved northwards as the population has declined, an extreme being reached in 1994, when sardine were only found north of the Cunene River, in Angolan waters. Likewise, there has been a major shift in the distribution of sardine in South Africa since the 1960s; the core of the distribution of adults having shifted from the West Coast to the South Coast. Acoustic surveys off Angola indicate that, in the 1990s, the biomass of both sardinella and Cunene horse mackerel has roughly doubled compared to the 1980s. Recent swept-area estimates of hake abundance in both Namibia and South Africa indicate that these populations are relatively stable at present, and there is some evidence that they could be gradually increasing. An interesting trend has been the increase in the abundance of Merluccius paradoxus in Namibian waters since 1992, which may indicate recent expansion or northward displacement of the M. paradoxus population from the South African West Coast. Acoustic and trawl survey estimates of adult horse mackerel in Namibia and South Africa over the past decade do not reveal any pronounced trends apart from a roughly 3-fold increase in the biomass on the South African West Coast since 1991. It is evident from CPUE indices that the abundance of line-caught species in South Africa has declined over the past few decades, to almost zero in some cases. As a result, the distributional ranges of many of these species has contracted, and the magnitude and extent of their migrations has declined. Aerial censuses reveal that the Namibian seal population was roughly halved by the effects of the 1993-1994 extreme anoxia events in the northern Benguela, from which the population now appears to have almost recovered. Aerial censuses have also revealed long-term reductions in the numbers of Cape gannets and Cape cormorants on the West Coast of southern Africa since the 1960s, and continuing declines in numbers of African penguins. A number of major effects of the environment on the distribution and abundance of commercially important resources in recent years are discussed. The most dramatic of these was the wide-scale advection of low-oxygen water into the northern Benguela from Angola in 1993 and 1994, and the subsequent Benguela Nińo of 1995, which appears to have severely affected the Namibian sardine population, and its major predators (particularly seals), and to have directly or indirectly increased mortality of juvenile hake on the Namibian shelf. The present socio-economic value of the region’s fisheries to Angola, Namibia and South Africa is outlined. Their national importance and the balance between the various sectors (industrial, artisanal, recreational etc.) varies considerably between the three countries. The fisheries sectors in Angola and Namibia rank high in national importance, for local food production in the case of Angola, and in Namibia for exports from the industrial fishery, which are worth over 1,35 billion N$ (approx. 225 million US$) per year at present. In both countries the sector is an important source of employment, which is largely informal in the case of Angola, and formal in Namibia. In South Africa, while the fishing industry is relatively unimportant nationally, exports rival those of Namibia, and the industry is an important source of revenue, food and employment in coastal areas, particularly in the Western Cape Province, which yields 80 - 90% of the total South African marine fish catch. Artisanal fisheries are currently of relatively little importance in South Africa, but the recreational fishery for linefish is large and varied, and directly or indirectly generates revenue and employment opportunities which probably exceed those of the industrial fishery. The broad national policies, legislation and formal structures for managing marine living resources in Angola, Namibia and South Africa are sketched, and the institutional capacity for research and management (including donor support) in each of the three countries is outlined. Current regional marine research programmes such as BENEFIT are briefly outlined, and other international and regional agreements pertaining to management of the region’s marine resources are listed. Recent and current methods used to assess the size of the major resources in each of the three countries are described in broad detail, and the methods used to manage them discussed. Major gaps in current knowledge of the region’s resources and threats to their rational management are discussed, with particular attention to trans-boundary issues, which are seen to be notable for hake, horse mackerel, sardine, anchovy and crab stocks. The major general scientific problems are seen as inadequate understanding of stock definition (particularly for shared or migratory stocks), inaccurate or non-existent information on basic biological characteristics (particularly age structure) for many of the harvested species, inadequate absolute estimates of population size and of trends in biomass, the lack of Operational Management Procedures based on population models for many of the resources, and the inability to predict the effects of environmental perturbations on resource dynamics with sufficient confidence for this information to be used in national or regional management. In most cases these problems are particularly severe in Angola and, to a lesser extent, in Namibia. It is noted that the root cause of many of the trans-boundary management problems is the lack of regional agreements and structures for research and management of shared resources, and the shortage of manpower and funds to undertake trans-boundary surveys and other related research activities. Finally, particular scientific and operational problems in Angola, Namibia and South Africa are discussed on a national basis. In Angola, where resources and their environment have been significantly less studied than elsewhere in the region, there are limited data series for retrospective analyses, large deficiencies in the understanding of fundamental life history characteristics of commercially important species, and no population models on which to evaluate management options. Catch and effort statistics are unreliable, and fisheries regulations are difficult to enforce owing to the extended coastline and small corps of compliance officials. Research capacity is limited because of the small number of people involved and the lack of appropriate local educational facilities. Most importantly, the breakdown of basic services and infrastucture as a result of the protracted civil war, and the severe macroeconomic problems in the country at present seriously inhibit development of local capacity in all spheres. In Namibia, the chief scientific challenges are the development of population models and Operational Management Procedures which would enable alternative harvesting strategies for the major fisheries to be formally evaluated, and the finding of objective ways of including environmental information into management decisions. A major operational constraint is the severe shortage of scientific and particularly technical staff within the Ministry of Fisheries and Marine Resources for the large number of resources which have to be studied, a problem exacerbated by the need for local staff to spend extended periods abroad for further education. In South Africa, the nation’s strong capacity in marine science is being threatened by a reduction in research funding, which has led inter alia to difficulties in maintaining even essential resource-monitoring surveys, and strict curtailment of environmentally-orientated cruises. Loss of senior research and management staff as a result of moves to reduce the size of the Public Service has resulted in a loss of experience and a greater load on remaining staff, which is likely to be aggravated by added responsibilities under the new Marine Living Resources Act. 
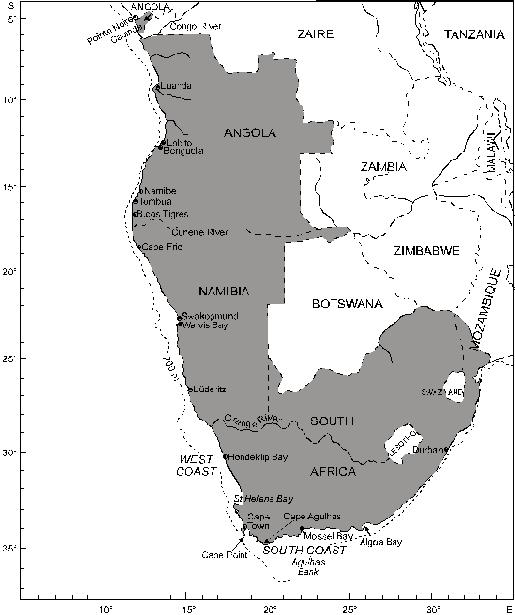
1. INTRODUCTION The Benguela Current Ecosystem can be loosely considered as covering the continental shelf between the Angola-Benguela frontal zone in northern Namibia/southern Angola and the Agulhas retroflection area, typically between 36 and 37 oS (Shannon and O’Toole 1998). As such, it covers the West Coast of South Africa, the entire Namibian coast, and southern Angola (Fig. 1) to an extent depending on the position of the Angola-Benguela front, which moves seasonally typically between 14 and 17 oS.
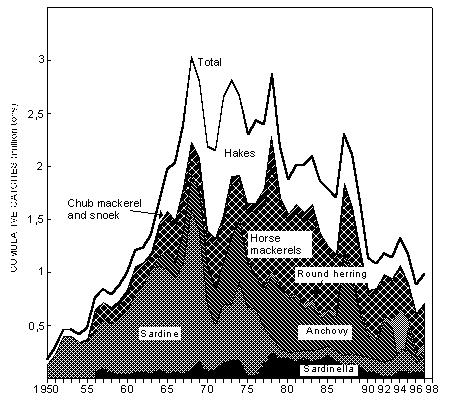
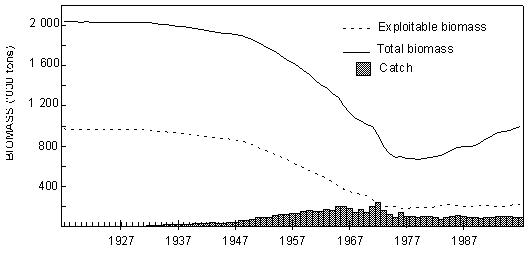
Fig. 1 Southern Africa, showing the 200m isobath, considered here to delimit the continental shelf The Benguela Current is one of the world’s major eastern-boundary current systems, and is rich in pelagic and demersal fish populations, supported by plankton production driven by intense coastal upwelling. These populations have been heavily exploited by man, particularly since the Second World War. Total fish catches in the south-east Atlantic climbed rapidly during the 1950s and 1960s, with the development of hake, sardine, anchovy, horse mackerel and sardinella fisheries (Fig. 2), and a valuable fishery for rock lobster in both Namibia and South Africa. The total annual catch peaked at over 3 million tonnes in 1968, but it subsequently declined to a level of around 2 million tonnes in the 1970s. This was largely attributable to major declines in sardine catches off both Namibia and South Africa, which were only partly compensated by increased (largely foreign) catches of hake and horse mackerel off Namibia. Total annual catches in the region subsequently dropped further to around 1.2 million tonnes in the 1990s, with a further sharp decline in catches of Namibian sardine in the second half of the 1970s, and the cessation of foreign trawling for hake and horse mackerel off Namibia after her independence in 1990. Since the 1960s there has also been a dramatic decrease in rock lobster catches, particularly off Namibia, where catches are now some two orders of magnitude below their peak in the 1960s. It is believed that most of these declines have been due to overfishing, although some of the major fluctuations have probably been influenced to a greater or lesser extent by the large-scale environmental perturbations that have occurred periodically in the system during this period (Shannon and O’Toole 1998).
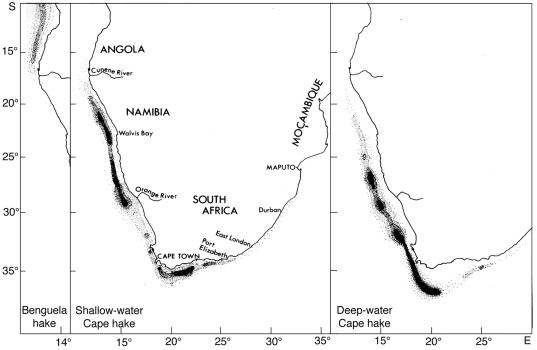
Fig. 2 Cumulative catches of the principal harvested species in the south-east Atlantic from 1950 This overview gives a brief description of the major living resources of the Benguela Current, and of the attempts that have been made, particularly in recent years, to manage them rationally and sustainably in each of the three countries bordering the system. It treats the fisheries regionally rather than nationally, and builds on material already presented in the regional Benguela Environment Fisheries Interaction and Training (BENEFIT) Programme Science Plan (Shannon and Hampton 1997), with particular emphasis on work carried out since the production of that document. References have largely been kept to recent key publications, and to a number of general articles from the region, wherein further references may be found. For the purposes of this overview, the Benguela ecosystem has been defined somewhat more widely than its physical limits, to include the extremities of the warm water systems which bound it on both sides. This is necessary, because some of the major resources of the Benguela spend a significant part of their lives outside the system boundaries, and are strongly influenced by interactions between the cool waters of the Benguela and the warm water of the Angola and Agulhas Currents. 2. OCCURENCE AND STOCK IDENTITY 2.1 Small pelagic fish The major fisheries for small pelagic fish off the west coast of southern Africa are those for sardine Sardinops sagax (also known as pilchard), anchovy Engraulis capensis, juvenile Cape and Cunene horse mackerel (Trachurus trachurus capensis and T. trecae respectively), round herring Etrumeus whiteheadi, and the round and flat sardinella (Sardinella aurita and S. maderensis respectively), which are fished almost exclusively by Angola. Sardine and anchovy live in temperate waters from southern Angola to KwaZulu-Natal in South Africa, with both species co-existing as quasi-discrete stocks off northern/central Namibia and off the Western Cape (i.e. between Cape Point and the Orange River). The degree of mixing between the southern and the northern populations of these species is unknown, but considering that the populations spawn in different, widely separated areas and are separated by a large, perennial area of cold, upwelled water off Lüderitz, it is probably not significant for management purposes, except in anomalous years. The round herring occurs over a similarly wide latitudinal range, but it appears to be most abundant east of Cape Point, particularly over the Central and Eastern Agulhas Bank. Most of the round herring caught in Namibian waters are juveniles, taken inshore as a small by-catch in the purse seine fishery. Although adults are occasionally taken farther offshore by bottom and midwater trawlers, the adult stock off Namibia is thought to be small compared to that farther south. Interaction between these stocks is probably of little consequence for management. Juveniles of Cape horse mackerel (i.e. fish < about 20 cm) are most commonly found off the west and south-west coasts of South Africa, and off northern Namibia/southern Angola, south of the Angola/Benguela front. These fish are believed to originate from separate spawning stocks off South Africa's South Coast and northern Namibia respectively. Juvenile Cunene horse mackerel T. trecae are found in subtropical and tropical waters in Angola and (occasionally) off northern Namibia. Their distribution extends from north-west Africa to the Angola/Benguela front, which moves seasonally between about 14 and 17 oS, with an average position at 16 oS (Pereira 1988, Shannon et al. 1987). Because of the highly dynamic nature of the front, it is most likely that the juvenile Cunene horse mackerel found in southern Angola, and very occasionally in northern Namibia, are part of a single population. Sardinella aurita and S. maderensis are found along the entire Angolan coastline, with the juveniles inshore, predominantly in the north. To the north their distribution extends apparently continuously along the coasts of Congo and Gabon, while to the south S. aurita can extend into northern Namibia in Benguela Nińo years. In the north both species undertake extensive spawning migrations along the Angolan coast (S. aurita more so than S. maderensis), making it unlikely that distinctly separate stocks exist in this region. It has however been suggested (Wysokiński 1986) that the Angolan stock of S. maderensis is independent of the stock off the coast of Gabon. Sardine tend to live within about 50 km of the coast, and are often found close inshore, both in South African and Namibian waters. Anchovy have a similar coastal distribution, but are commonly found more than 100 km offshore on the Agulhas Bank off the Cape South Coast in the spawning season. Round herring are found widely distributed across the shelf both on the West Coast and the Cape South Coast, with a clear increase in size with distance offshore. Juvenile Cape horse mackerel off both South Africa and Namibia, and juvenile Cunene horse mackerel off Angola, are most abundant inshore, generally being found within the 100m isobath throughout the region. S. aurita inhabits the continental shelf and is generally found in calm saline waters (>35 ppt) at temperatures around 24 oC. In contrast, S. maderensis is a coastal, more euryhaline species, also generally found at temperatures above 24 oC, often in the vicinity of river mouths (Luyeye 1995). The two species appear to be roughly equal in abundance, except in the south where S. aurita predominates. Surveys indicate that sardinella density is generally higher in the central (Luanda-Benguela) region than to the north and south (Luyeye 1995). 2.2 Trawled fish The major species caught by trawl off Namibia and South Africa are the Cape hakes Merluccius capensis and M. paradoxus, which are caught in bottom trawls, and adult Cape horse mackerel, which are mostly caught in midwater trawls off Namibia and in bottom trawls off South Africa as a by-catch in the hake fishery. Other significant by-catch species in the hake fishery in both Namibia and South Africa are monkfish Lophius spp, kingklip Genypterus capensis, snoek Thyrsites atun and the West Coast sole Austroglossus microlepis. In recent years, the monk fishery in Namibia has become increasingly directed, with the West Coast sole as the most important by-catch. On the outer Namibian shelf there is also a valuable deep-water trawl fishery directed at orange roughy Hoplostethus atlanticus and, to a lesser extent, alphonsino Beryx splendens and other deep-water species. Off Angola there is a relatively small bottom trawl fishery for Benguela hake Merluccius polli and M. capensis (in the extreme south), and more important ones in central and northern Angola for demersal species such as Dentex spp. and red pandora Pagellus belloti. The large-eye dentex Dentex macrophthalmus is also taken off northern Namibia in midwater trawls, together with jacopever Helicolenus dactylopterus, another important by-catch species. The distributions of the three species of hake in the Benguela region are shown in Figure 3. Merluccius polli occurs predominantly in Angolan waters, and is caught on the shelf slope as a by-catch in the prawn fishery and by deep water trawlers in the south, where its distribution overlaps with that of the shallow-water Cape hake M. capensis. The two species of Cape hake are found throughout Namibian and South African waters. M. paradoxus (deep-water hake) occurs in deeper water than M. capensis, although the two species co-occur at intermediate depths (Payne 1989). Typically the former is found in water 150 - 800m deep, mostly at temperatures between 4 and 8oC, whereas the latter occurs from the coast to a water depth of about 380m, in temperatures between 4 and 12 oC. It has been suggested (Payne 1989) that, in South Africa, the relative abundance of the two species is related to the width of the shelf and the steepness of the slope, with M. capensis predominating where the shelf is broad and the slope steep, and vice versa for M. paradoxus. Larger individuals of both species are found at greater depths than smaller fish, and there is little overlap in the distribution of mature fish. M. capensis is the more common species off Namibia, especially in the central region, although M. paradoxus has been increasingly abundant and more widely distributed there in recent years (see following). M. paradoxus predominates off the west coast of South Africa. It is believed that this stock may be the origin of the Namibian M. paradoxus stock. A second population of M. capensis, which for management purposes is treated as a separate stock, exists in the extreme southern Benguela, chiefly over the Agulhas Bank In the 1970s and 1980s, ICSEAF treated Cape stocks off southern Angola/northern and central Namibia (15 - 25oS), southern Namibia (25 - 30oS), the South African West Coast (30oS - 20oE), and the South African South Coast as separate for management purposes. The species were considered together, as the catch records did not distinguish between them Fig. 3 suggests that the West Coast stocks of both species are probably shared between Namibia and South Africa, although catch patterns between Lüderitz and the Orange River indicate that there may be a measure of separation between the Namibian and South African M. capensis stocks. In contrast, there is some evidence from surveys (e.g. Strřmme 1996) and commercial catches that, since 1990, there has been a gradual migration or expansion of M. paradoxus into southern Namibia and farther north, probably from South African waters. This is the only reported evidence of significant longshore movement of any of the three hake species, although it has previously been suggested that such shifts do occur in response to environmental changes such as temperature fluctuations and the movement of de-oxygenated bottom water.
Fig. 3 Distribution of the three hake species in the Benguela ecosystem (from Payne1989) At least two stocks of Cape horse mackerel exist off southern Africa, viz. off northern Namibia/southern Angola, and off the Western Cape. These stocks were once believed to be genetically separated by the environmental barrier of the Lüderitz upwelling cell, with only a limited interchange between them. However, the finding of large adults on the bottom across the shelf south of Walvis Bay in recent Namibian surveys (E. Klingelhoeffer, NatMIRC, pers. comm.) raises the question of a possible connection between this part of the population and that on the West Coast of South Africa. This is a regional question of major importance for managing this trans-boundary fishery. Opinion is divided on whether the horse mackerel on the Cape West and South Coasts form a single or separate stocks, with catch patterns suggesting the former and genetic studies the latter. There is little information regarding the stock structure of Cunene horse mackerel in Angolan waters, although Sardinha (1996) has suggested on the basis of biological data and distribution patterns that there are separate self-sustaining populations in the north and south of Angola. The hypothesis is currently being tested genetically (Sardinha and Nćvdal, submitted). Orange roughy in Namibia are found mainly over the shelf between about 600m and 1000m depth at bottom temperatures of between 3 and 7 oC (A. Staby, NatMIRC, pers. comm.). The fish tend to be concentrated over hard substrata in a number of small areas, particularly during the spawning season. Alfonsino tend to be more widely distributed over the outer shelf, between about 400 and 700m. The degree to which the distribution of the two species extends into Angolan and South African waters, and the extent of any longshore migrations, is at present unknown. The main commercial species of monkfish found in southern African waters are Lophius vomerinus (previously known as Lophius upsicephalus) and Lophius vaillanti. The former is found from northern Namibia to the East Coast of South Africa, but the latter only north of Walvis Bay. Both are demersal species, mainly found at depths of between 150 and 400m. Two separate recruitment areas for L. vomerinus have been located in Namibia: off Walvis Bay and near the Orange River. The relationship between these recruits and L. vomerinus to the south is unknown, as is the extent of any longshore migrations of adults. Nonetheless, it seems reasonable to assume that there is some interaction between the population(s) on the South African West Coast, and that found around the Orange River, in particular. Demersal fish caught commercially by trawl in Angola can be grouped into species occurring below the thermocline along the continental shelf, and species found above, below and in the thermocline. Sparids are the most important in the former group. particularly the large eye dentex Dentex macrophthalmus, which is fished between 60 and 300m depth between Lobito and as far south as Walvis Bay (Constança 1995). Other important species are the Angola dentex Dentex angolensis and the red pandora Pagellus belloti, which occur across the shelf to depths of around 300m (Bianchi 1986). These three species together usually make up more than half of the demersal catch. In the latter group the most important species is the bigeye grunt Brachideuterus auritus. A significant part of the demersal trawl catch consists of deep-water prawns and to a lesser extent, deep-water red crab, which are discussed in the following Section. 2.3 Crustaceans The major crustacean fisheries along the west coast of southern Africa are those for the West Coast rock lobster Jasus lalandii off South Africa and Namibia, the red crab Chaceon maritae off northern Namibia and Angola, and for the deep-water rose prawn Parapenaeus longirostris and striped red prawn Aristeus varidens off northern and central Angola. J. lalandii is a spiny lobster associated with the cool upwelled waters of the Benguela. It occurs in commercially exploitable densities from east of Cape Point to approximately 25oS, and at lower densities beyond its core distribution. Close inshore it is caught by hoopnets deployed from dinghies and by recreational divers, but in deeper waters is harvested by traps. C. maritae occurs on the slope of the continental shelf from about 27 oS off Namibia, northwards to Angola, Congo and the Ivory Coast. Off Namibia it is found on soft mud substrata at depths of between about 300 and 900m, and is harvested solely by Japanese vessels using traps. Off Angola it is found within a similar depth range, particularly in the southernmost area, and is taken in traps and (occasionally) bottom trawls. It has recently been shown from tagging studies (Le Roux 1997) that adult females migrate from Namibia to Angola, suggesting a single stock in the region, which needs to be managed jointly by Namibia and Angola. Parapenaeus longirostris is essentially an Atlantic Ocean prawn, found from Portugal to Angola in the east, and from Massachusetts, USA, to French Guiana in the west, whereas Aristeus varidens is an East Atlantic species, found from Rio de Oro (24 oN) to 18 oS (off Namibia). The depth disrtribution of the two species differ. P. longirostris is found on the continental shelf and upper slope, between 50 and 400m depth (López Abellán and de Cárdenas 1990) over sandy bottoms. A. varidens lives on the slope, mainly between 400 and 800m depth, and is strongly associated with muddy bottoms. The size of both species increases with depth, more so in the case of P. longirostris than A. varidens. Angolan-Spanish surveys have revealed that P. longirostris have a more homogeneous distribution than A. varidens, which tends to concentrate in several areas, mainly related to submarine canyons. Concentrations of P. longirostris tend to occur around recruitment areas. 2.4 Line-caught species The linefisheries of the Benguela Current and adjacent waters exploit a large number of species. They can be broadly classified into a) inshore reef fishes, which are mainly resident on shallow nearshore reefs and have a limited geographic distribution, b) migratory shoaling species, where the adults aggregate and migrate rapidly over large distances, usually as part of annual migratory cycles, and c) offshore large pelagic species such as tuna and billfish, which form large, highly-migratory straddling stocks that cross the borders of many countries, and even oceans. In Angola, line-caught species belonging to the first two groups make up more than 40% of the total catch by the large inshore artisanal/subsistence fishery which extends along the entire coast. The most important groups caught by line are dentex, croakers (Sciaenidae) and groupers (Serranidae). Catches are highest in the provinces of Benguela, Namibe and Luanda (Anon. 1998a). The recreational fishery for linefish in Angola is underdeveloped and negligible compared to the artisanal/subsistence fishery. The silver kob Argyrosomus inodorus is the most important of the linefish species caught commercially in Namibia, with roughly equal amounts being taken by commercial fishermen and recreational anglers. Other important angling species in Namibia are the steenbras Lithognathus aureti, the blacktail Diplodus sargus and the galjoen Coracinus capensis. Off the South African West Coast, the dominant reef fish in the catches is the hottentot Pachymetopon blochii, which has been an important contributor to artisanal/subsistence linefish catches off the West Coast since the early part of the century. It is a highly resident species, which makes it susceptible to localised depletion in heavily fished areas. In contrast, the galjoen Coracinus capensis, which is a major shore-angling species off Namibia and the West Coast of South Africa, is highly migratory. Tagged galjoen have been found to have moved from northern Namibia to east of Cape Point. Snoek is by far the most important migratory linefish species caught commercially on the West Coast of South Africa, and is important in Namibia as well. It is found along the entire southern African coast from southern Angola to Cape Agulhas, mainly in cool upwelled water, and is a major predator on pelagic fish in the region. Snoek were historically considered to form a single stock extending from Cape Agulhas to northern Namibia, and to migrate seasonally between these regions (e.g. Crawford et al. 1990). However, a recent study of all available evidence (Griffiths, in prep.) suggests that there may be two separate sub-populations - in Namibia and South Africa respectively - with medium-term (of the order of five years) exchange between them in response to environmental events and food availability. Of the large pelagic species taken in the region, the most important in the southern part is the albacore or longfin tuna Thunnus alalunga, which is currently caught by South African and Namibian pole and line vessels within territorial waters from south of Cape Point to Lüderitz. The species is also exploited by Asian high-seas longliners off both countries. The stock is believed to be part of a single southern Atlantic stock, separated from the southern Indian Ocean stock by the warm water of the Agulhas current. There is also a longline fishery for bigeye tuna Thunnus obesus along the edge of the shelf in both countries, mostly by Asian high-seas vessels. It is not known whether these fish form part of an Indian Ocean, Atlantic Ocean or circumglobal stock. In Angola the most important species taken by local baitboats is the yellowfin tuna Thunnus albacares, while bigeye tuna is the major constituent of the Japanese longline fishery. Yellowfin form part of an Atlantic population, which spawn off Brazil and the Gulf of Guinea, and are most abundant in southern Angola in summer. Finally, mention must be made of the Cape hake Merluccius capensis, which although primarily a trawl-caught species, is also caught by lines off South Africa's South East Cape, and (to a lesser extent) Namibia, using both hand and hydraulically-hauled longlines. The South African fishery grew out of an experimental longline fishery for kingklip in the 1980s, which severely depleted that stock. 2.5 Seals and seabirds The Cape fur seal Arctocephalus pusillus pusillus occurs along the southern African coast between Algoa Bay and southern Angola and is harvested in Namibia. Although the harvest is low compared to earlier times (the fishery is centuries old), seals have been included in this overview because they are major top-predators in the Benguela, whose dynamics have been strongly affected by fluctuations in a number of the major fish resources of the region, making them important visible indicators of environmental change. The same is true of resident seabirds such as the Cape gannet Morus capensis, the Cape cormorant Phalacocorax capensis and the African penguin Spheniscus demersus, which breed mainly on nearshore islands and guano platforms off Namibia and South Africa, and feed largely on pelagic fish such as sardine and anchovy. The dynamics of pelagic fish stocks is strongly reflected in changes in abundance, diet and breeding success of the seabirds, to the extent that in South Africa, consideration is being given to using this information directly in the management of the sardine and anchovy fisheries. 3. LIFE HISTORY OF MAJOR RESOURCES 3.1 Small pelagic fish Sardine and anchovy Off Namibia, sardine spawn largely within 60 km of the coast in two main areas, one off Walvis Bay and the other farther north, in the mixing zone south of the confluence of the Benguela and the Angola Current systems. In the northern area, peak spawning (mainly by young adults) occurs near the 200m isobath in late summer/autumn in water temperatures between 19 and 21 oC, whereas spawning farther south (mainly by older fish) takes place in summer in cooler water close to upwelling zones. Since the collapse of the sardine stock in the 1970s (Section 4.1), spawning in the south has diminished in importance. The distribution and movement of anchovy off Namibia is similar to that of sardine there, but significant spawning only occurs north of Walvis Bay. The larvae of both species drift south close to the coast, recruiting as 0-group fish into the fishery in the cool upwelling areas near Walvis Bay. This is followed by a return northward migration of juveniles and young adults to the northern mixing area, where they first spawn. In the case of sardine, older fish subsequently return south again to spawn in the Walvis Bay region, although this migration is believed to have decreased in importance since the collapse of the fishery. The behaviour of sardine and anchovy off Namibia is analagous to that found in other upwelling systems, such as those off California, Peru and north-west Africa (Bakun 1995), where both spawning and recruitment occur downstream of the principal upwelling cell (Lüderitz in this case). In the past, sardine in the southern Benguela have spawned in two areas, one of large adults in cool water west of Cape Columbine, and the other of younger fish in warmer water east of Cape Point, inshore of the Agulhas Current, analagous to the two sardine spawning areas off Namibia. However, since the major stock decline in the mid-1960s, there has been little evidence of spawning on the West Coast, and it seems that the stock is now perpetuated mainly by the younger adults spawning over the Agulhas Bank, particularly the Western Bank, between Cape Point and Cape Agulhas. Anchovy also spawn mainly over the Agulhas Bank, somewhat offshore of the sardine, in the upper mixed layer, in water temperatures ranging between 17 and 19 oC. Peak anchovy spawning is in early to mid-summer, whereas sardine spawn over an extended period, with weak maxima during late winter/early spring and late summer. In contrast to the northern Benguela, spawning occurs mainly upstream of the main upwelling centres of Cape Point and Cape Columbine. Eggs and larvae are transported from the Western Agulhas Bank to the West Coast (Fig. 4) between the 200m and 500m isobaths (Fowler and Boyd 1998), along the offshore boundaries of the upwelling cells by a perennial equatorward jet current, leading to recruitment downstream of the upwelling cells. Bakun (1995) has noted that this is a reversal of the normal spawning and recruitment pattern in other eastern-boundary upwelling areas (e.g. the northern Benguela, California Current and Canary Current), the difference originating from the right-angular configuration of the coastline in the southern Benguela
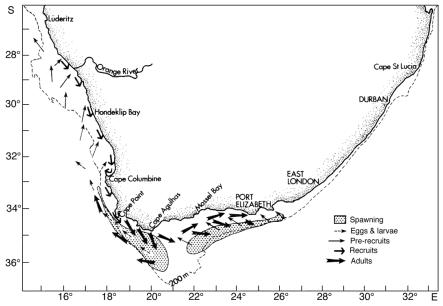
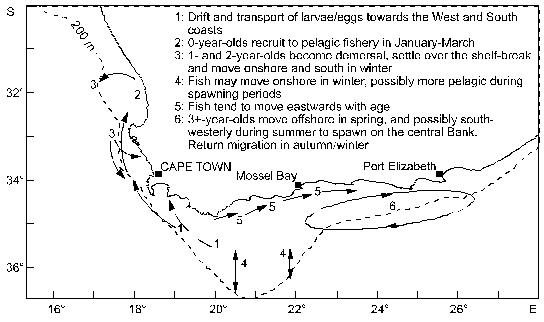
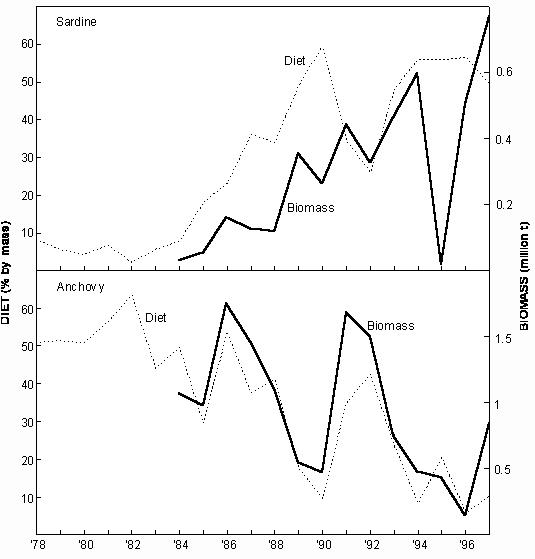
Fig. 4 Conceptual model of anchovy migration, constructed from acoustic survey data and trends in length in research midwater trawl catches (from Hampton 1992) Sardine and anchovy larvae transported past Cape Columbine are either carried offshore and lost to the system, or are transported northwards and inshore (possibly assisted by active swimming as they develop), leading to recruitment of both sardine and anchovy inshore along the West Coast at least as far north as the Orange River (Fig. 4). There is evidence that in normal years, the northern limit of the dispersal is the southern edge of the Lüderitz upwelling cell, which causes the larvae either to be carried far offshore, or to be drawn inshore by compensatory flow and returned south by the inshore poleward counter-current. It has been suggested that in years when this cell is abnormally weak (as in 1987), pelagic larvae spawned in Cape waters may be carried past the Lüderitz "barrier" and recruit into the southern Namibian fishery, linking the South African and Namibian pelagic fisheries. This may have occurred in 1987, when an unexpectedly large number of anchovy recruits was caught in southern Namibia, resulting in catches roughly an order of magnitude higher than in the previous and following years. Once inshore, the sardine and anchovy recruits off South Africa move south close inshore along the West Coast in autumn and winter, assisted by the poleward counter-current. They are possibly retained for a period in St Helena Bay before reaching the Agulhas Bank in early summer, towards the end of their first year of life. It is during this period that they are exploited by the purse-seine fleet, which mainly operates from ports on the West Coast. Adults tend to move eastward and (in the case of anchovy) offshore, with increasing age, although there is some return of older fish to the West Coast from the outer edge of the Agulhas Bank, probably assisted by the jet current off the Cape Peninsula. A portion of the sardine population (mostly fish in their second year of life) undertakes a pronounced and well-documented inshore migration into the shelf waters of KwaZulu-Natal each winter (known locally as the "Sardine Run"), to at least as far up the coast as Durban. This is probably in response to the eastward retreat of warm subtropical water close inshore at that time of the year, and is assisted by equatorward counter-currents inshore of the Agulhas Current. The migration may be analagous to the movement of young Namibian sardine towards the subtropical system boundary in winter. It is most probable (though not yet demonstrated) that the surviving fish return rapidly to the Agulhas Bank later in the year in the strongly flowing Agulhas Current slightly farther offshore. Analyses of stomach contents up to the end of the 1970s suggested that the diets of sardine and anchovy in the Benguela are similar, with phytoplankton as the main food source. However, laboratory and field studies since then have shown that zooplankton is more important in the diet of both species than was previously believed to be the case. Certainly, juvenile sardine and anchovy, and adult anchovy, feed primarily on zooplankton, although adult sardine appear to utilise more phytoplankton in areas of consistently high phytoplankton abundance. Little is known about the extent to which sardine and anchovy compete for habitat and food in the Benguela ecosystem, but a recent field study on feeding in mixed schools of juveniles, in which a clear size difference was found in the zooplankton taken by the two species (Louw et al. 1998), suggests that direct competition may be limited, at least in the younger stages. Sardinellas The main spawning area of S. aurita and S. madarensis is thought to be between 5 and 7 oS (Pointe Noire to south of the Congo River), with peak spawning in March-April. There appears to be a seasonal longshore migration pattern for both species, with an equatorward movement towards the spawning grounds during the first part of the year, and a return poleward migration of adults in the second half of the year to central and (in the case of S. aurita) southern Angola. Juvenile S. aurita are encountered over the whole littoral zone from Cape Lopez in Gabon to Baia dos Tigres in southern Angola. Upon reaching a length of 10 – 14 cm, the juveniles leave the littoral zone, and remain for some time in shallower shelf waters before joining the adult stock farther offshore (Troadec and Garcia 1980). Juvenile S. maderensis appear to be largely concentrated inshore north of the Congo River throughout the year (from data in Wysokiński 1986). Little is known about the diet of sardinellas in Angola, although French studies in the 1970s found that the diet of both S. madarensis and S. aurita in Congolese waters consisted almost entirely of the copepod Calanoides carinatus. Round herring Little is known about the seasonality of round herring spawning in the Benguela, although from ichthyoplankton surveys in the south, it appears that there at least, spawning probably occurs throughout the year, reaching a peak between late winter and early summer. The early life history of round herring spawned on the Agulhas Bank appears to be similar to that of sardine and anchovy spawned there, with the ichthyoplankton being transported to the West Coast by the same jet current, leading to recruitment along the West Coast at the same time of the year as sardine and anchovy recruitment, and movement onto the Agulhas Bank towards the end of the first year of life. The fish appear to move eastwards and offshore with increasing age, inhabiting the entire Agulhas Bank, at least as far east as Port Elizabeth (Roel and Armstrong 1991). As with sardine and anchovy, there is probably some return of older fish to the West Coast from the outer regions of the Bank. Round herring is a particulate feeder, whose diet in South African waters consists exclusively of zooplankton (mainly copepods, euphausiids and decapods). Juveniles form pelagic shoals in the upper mixed layer, but adults perform pronounced diel vertical migrations, migrating from near the surface at night to near-bottom waters during the day, often passing through a temperature gradient of more than 10 oC. 3.2 Trawled species Hakes Cape hakes spawn in midwater throughout the year, with a peak in early summer for both M. capensis and M. paradoxus, and a secondary peak in late summer for M. paradoxus in the southern Benguela. Most M. paradoxus spawning is thought to take place along the edge of the Agulhas Bank, but spawning also occurs over the shelf-break west of St Helena Bay and off central Namibia. In the latter region, M. capensis spawn most frequently between 160 and 250m depth, spawning starting earliest in the shallower waters. The eggs of both species are concentrated around the depth of the thermocline, and the dispersal of M. paradoxus eggs and larvae produced on the South Coast could be similar to that of the pelagic fish spawning there, resulting in young M. paradoxus being plentiful between Cape Columbine and the Orange River. To the north, 0-group M. capensis are particularly abundant off Walvis Bay, which appears to be a nursery area. Juveniles of this species are also plentiful off the Orange River and south to about Cape Columbine, sometimes co-occuring with pelagic fish recruits in winter, on which they feed (particularly on anchovy). As with the pelagic recruits, juvenile hake in the Orange River area move south as they grow older, but unlike them they tend to move offshore as they move south. There is evidence that off the West Coast of South Africa, juvenile M. paradoxus move inshore in summer and offshore in winter, in response to changing feeding regimes. The adults, which tend to concentrate in depths greater than about 500m in summer and autumn, move inshore in spring to depths of around 300m, and then return offshore, movements which are probably related to both spawning and feeding. Off Namibia, M. capensis follows a similar inshore migration during the spawning season in early summer, followed by an offshore migration in late summer. Cape hakes feed both close to the bottom and in midwater, and tend to be off the bottom at night, although no clear feeding periodicity in either M. capensis or M. paradoxus has been demonstrated, except in the case of juvenile M. capensis, which off the West Coast of South Africa have been observed to move off the bottom at night to feed on pelagic prey such as juvenile anchovy, and to return to the bottom before dawn (Pillar and Barange 1995). Recent studies (Pillar and Barange 1998) have indicated that M. capensis adults on the South African West Coast also move into midwater at night in response to the vertical migration of their prey, but that they only return to the bottom when satiated, regardless of time of day. This results in aperiodic, asynchronous vertical movements of individuals, depending on food availability and recent feeding activity. This lack of a distinct diel feeding rhythm has also recently been reported by Huse et al. (1998) from studies on the behaviour of M. capensis and M. paradoxus at a location on the central Namibian shelf, although they did find some evidence of increased feeding in the early evening, in common with earlier studies of M. capensis feeding in the same area. Cape hakes are opportunistic feeders, resulting in considerable seasonal and spatial variability in their diet. On the South African West Coast, young M. capensis and M. paradoxus feed predominantly on planktonic crustaceans (particularly euphausiids), juvenile anchovy (by M. capensis), lightfish Maurolicus muelleri and lanternfish Lampanyctodes hectoris (by M. paradoxus), the diet of both species becoming increasingly piscivorous with age (Punt et al. 1992). Squid, epipelagic fish and, to a lesser extent, mesopelagic fish such as lightfish and lanternfish comprise a significant proportion of the diet of adult M. capensis, but in the larger fish the principal diet items are small M. paradoxus, small M. capensis (to a lesser extent) and other demersal species (Punt et al. 1992). With increasing age, M. paradoxus becomes increasingly cannibalistic on young M. paradoxus, which together with squid, crustaceans and mesopelagic fish constitutes most of the diet of the large mature adults on the Cape West Coast. In contrast, more than 90% of the diet of large M. capensis on the Agulhas Bank consists of pelagic fish (particularly anchovy), horse mackerel and young M. capensis, the size of prey increasing with hake size (Pillar and Wilkinson 1995). Cannibalism on M. capensis appears to be higher than on the West Coast, but the interspecific (hake-on-hake) predation somewhat lower. The diet of hake in Namibia is similar to that in South Africa (see summary in Gordoa et al. 1995), the chief difference being the greater importance of myctophids (particularly for M. paradoxus), and the significant proportion of gobies Sufflogobius bibartus in the diet of both species. As in South Africa, both species become increasingly piscivorous with age, and hake-on-hake predation becomes increasingly important. Because of their catholic feeding habits and abundance, hake are extremely important predators in the Benguela. For example, Punt et al. (1992) have estimated that Cape hakes in South African waters could consume as much as 6 million tonnes of food annually. Based on estimates of stock size in Namibia, it would appear that consumption there could be as high. Horse mackerels Cape horse mackerel T. trachurus capensis off the West Coast of southern Africa have broad spawning areas, with most intense spawning in warmer waters immediately west of the shelf-break in both regions. Egg and larva surveys in the 1970s (O’Toole 1977) showed that in Namibia the heaviest spawning occurs in the north between October and March in the mixing zone of warm oceanic water and cool coastal water, and that the timing of spawning is closely linked with the duration and intensity of mixing. Nursery areas exist in both the southern and the northern parts of the ecosystem, adjacent to the spawning grounds but closer inshore, and there are substantial longshore and cross-shelf migrations of both juveniles and adults. Off Namibia, juvenile Cape horse mackerel live inshore, the smallest fish being found farthest north. Slightly larger individuals appear to migrate south towards Walvis Bay, especially in winter. Maturing fish move offshore and northwards to spawn, the adults generally occuring north of 21 oS. Off the Western Cape, juveniles also occur inshore and the adults farther offshore. The movements of the fish with increasing age, and particularly the interchange between the West and South Coast populations, are particularly complex, as can be seen from a recent migration model postulated by Barange et al. (1998), reproduced here as Fig. 5. Note that the life history in the first year of life is similar to that of sardine and anchovy, with spawning on the South Coast leading to recruitment on the West Coast in the following year, probably driven by the same transport mechanisms. Recruitment occurs about three months earlier than that of sardine and anchovy, but overlaps with it to some extent, resulting in a by-catch of juvenile horse mackerel in the pelagic fishery. Thereafter the species diverge, with the horse mackerel becoming more demersal and moving offshore, probably ultimately leading to spawning over the shelf-break on the West Coast. Some of these fish however move onshore again in winter in their second year of life, and move onto the Western Agulhas Bank, assisted by the poleward counter-current on the inner shelf. These fish reach maturity at two years of age and move eastwards and offshore with increasing age, leading to spawning across the entire Agulhas Bank, which peaks at different times on the Western and Eastern Banks and which, on the Western Bank at least, appears to be closer inshore in winter than in summer. Barange et al.(1998) suggest that adults on the Eastern Bank move southward and westward to spawn on the Central Bank in spring, a movement which would favour transport of the ichthyoplankton to the West Coast. They also suggest that there is some secondary recruitment directly into the numerous bays of the South Coast from spawning farther offshore, probably aided by the inshore movement of spawners in winter. However, most of these hypotheses are still tentative and need rigorous testing. Fig. 5 Conceptual model of Cape horse mackerel migration patterns in the southern Benguela, constructed from acoustic and research trawl information (from Barange et al. 1998) According to Troadec and Garcia (1980), the core spawning area of the Cunene horse mackerel is off Mauritania and Senegal, where spawning peaks from February to June. Although spawning undoubtedly occurs farther south as well, including in Angolan waters, little has been reported about the locality and timing of any such spawning. Little is also known about the seasonal distributional patterns of the different life history stages of the species off Angola, although the fact that at the height of the fishery in the 1960s and 1970s, inshore catches used to show a sharp peak in October/November co-incident with a rise in water temperature, has led to suggestions that there is a seasonal inshore migration in response to changing environmental conditions. This is supported by survey data, which indicate that the proportion of the population in central Angola is highest in summer (Sardinha 1996). Cape horse mackerel up to the age of two years feed near the surface and are planktivorous. The diet, which consists mainly of copepods, is similar to that of sardine and anchovy, and juveniles up to about 10 cm in length can co-exist in schools with sardine and anchovy. Adults off the west coast of southern Africa are opportunistic feeders on euphausiids (which constitute 95% of their diet in Namibia), polychaete worms, chaetognaths, squid, various crustaceans and fish such as gobies Sufflogobius bibartus, lanternfish and lightfish. Older horse mackerel tend to feed in midwater, and their diet is similar to that of Cape hakes of similar size. Accordingly, there may be interspecific interaction between Cape horse mackerel and Cape hakes, with a decrease in the abundance of the one species benefiting the other, and vice versa. Pillar and Barange (1998) showed that adults on the Cape South Coast feed predominantly on copepods close to the bottom in late afternoon, migrate in synchrony into midwater at dusk, clearly for purposes other than for feeding, and return to near-bottom at dawn. Sardinha (1996) reports that in Angolan waters, horse mackerel of both species undergo a similar diel vertical migration. There have been no comparable studies in Namibian waters, but it would appear from the fact that adult horse mackerel there are caught by midwater trawl throughout the 24-hour period that their vertical migratory behaviour in this region may be different from that to the north and south. Deep-water species Orange roughy off Namibia have a short spawning period of about a month in July/August, when they spawn in dense concentrations close to the bottom in small areas typically no more than 10 - 100 km2 in extent. They are exceptionally long-lived and slow-growing, possibly only reaching sexual maturity at around 25 years off Namibia, and may have a maximum lifespan of over 100 years. The fish have a low reproductive rate, which together with their aggregating behaviour, makes them highly vulnerable to over-fishing. Alfonsino Berxy splendens are distributed over a wider area and are probably more productive. Little is known about their spawning behaviour or breeding habitat. Dentex Dentex macrophthalmus spawn throughout the year with a peak between October and April. The most intensive spawning occurs between Baia dos Tigres and the Cunene River at depths between 25 and 110m (Kuderskaya 1985). Little is known about migration patterns, except that according to Wysokiński (1986), the stock migrates shorewards in summer and offshore in winter. Even less has been reported on the life histories of the other commercially exploited demersal fish in Angolan waters. 3.3 Crustaceans West Coast rock lobster J. lalandii has a well-defined moulting and spawning cycle. Adults moult once per year, the males in spring and the females in late autumn/winter, after which mating takes place. Egg-hatching peaks in October-November and the phyllosoma larvae remain planktonic for a long period, drifting in oceanic sub-gyres until they reach the puerulus (free-swimming) stage and settle. Females reach sexual maturity about five years after settlement, at a greater length in the south than in the north. Maturing males grow faster than females, resulting in the fishery being based largely on males. The adults are generally distributed offshore of the juveniles, except in the north, where the population is constrained close to the coast by low-oxygen water. J. lalandii feed largely on mussels, in particular the ribbed mussel Aulacomya ater, which is abundant in the rocky subtidal zone of the West Coast. In areas of low mussel abundance, the diet consists mainly of echinoderms (sea urchins and starfish), gastropods, bryozoans, polychaetes and seaweeds. The principal predators on J. lalandii are octopus, dogsharks, hagfish, whelks (on injured or weakened animals) and young seals. Cannibalism is known to be prevalent in overcrowded situations, particularly among juveniles. Deep-sea red crab Chaceon maritae in Namibia appear to spawn throughout the year, judging from the fact that no seasonal cycles in moulting and egg-bearing have been found there (Le Roux 1997). Adult females generally live in shallower water than males, and virtually all egg production and larval release takes place on the shallower part of the continental slope; a pattern which has also been found in Angola. The fact that the migration from Namibia to Angola is almost entirely by females suggests that this is a spawning migration. Deep-sea prawns Parapenaeus longirostris spawn throughout the year, with peaks in July and December. Sobrino and de Cardenas (1989) state that females reach sexual maturity at a carapace length of 21.6 mm. According to López Abellán and Garcia-Talavera (1992), there are nursery areas between 8 and 9 oS and between 10 and 11 oS. Eggs are demersal and the larvae planktonic. The larvae enter the post-larval phase at a length of 12 mm. Juveniles are concentrated between depths of 50 and 70m, where recruitment takes place. A sampling programme has shown that there is a major recruitment peak between January and March, and a lesser one in August-September (López Abellán et al. 1993). Aristeus varidens appear to spawn throughout the year, with a peak in December and probably another in April-May. First maturity is reached at a carapace length of about 25 mm (Sobrino and de Cárdenas). 3.4 Line-caught species Snoek spawn along the edge of the shelf off the West Coast of South Africa and the western Agulhas Bank, and off southern Namibia, mainly from July to October. There is also some evidence of spawning farther north. It has been suggested that in South African waters the ichthyoplankton is transported by prevailing currents to a primary nursery ground north of Cape Columbine, and to a secondary one east of Danger Point on the South-West Coast (Griffiths, submitted). Juveniles tend to recruit inshore and remain as locally migratory shoals in nearshore nursery areas until they approach maturity, when they join the adult population. Their cross-shelf distribution on the inner shelf is determined by prey availability, and includes a seasonal inshore migration in autumn in response to pelagic fish migration patterns. Adult snoek are found throughout their distributional range, moving offshore and somewhat southwards to spawn. Other than this, there do not appear to be any seasonal trends in longshore movements of adults in South African waters (Griffiths, in prep.). Migrational patterns of juveniles and adults in Namibian waters have not been established with any certainty. Albacore, the main contributor to the South African and Namibian large pelagic fisheries at present, are believed to migrate across the southern Atlantic to South America, and then northwards to spawn in the tropical central Atlantic. Juveniles occasionally recruit into waters off the Western Cape, but most of the fish caught are large reproductively inactive adults, following and feeding on the rich pelagic prey in the Benguela and Agulhas Current systems. 3.5 Seals and seabirds The Cape fur seal breeds on small rocky nearshore islands and, most importantly, at six mainland colonies on the Namibian and northern Cape coasts where human access is restricted. Two of these colonies (Kleinsee in the northern Cape and Atlas Bay near Lüderitz) are believed to be the largest mainland seal colonies in the world. The breeding season, during which pupping is followed almost immediately by mating, lasts for 6 to 8 weeks in October/November. Pups are weaned at an age of 8 to 10 months, and thereafter forage widely. While attending their pups, adult cows feed within a few days range of the colonies, but the bulls appear to have separate feeding grounds, probably considerably further offshore. Much of the diet is made up of fish, of which commercial pelagic fish and hake are the most important on the South African West coast, and the bearded goby Sufflogobius bibartus (a non-commercial species), horse mackerel and juvenile hake the most important off Namibia. It has been estimated that seals in the Benguela consume about 1 million tonnes of fish annually, which is of the same order as the total annual fish catch in Namibia and South Africa. The Cape gannet breeds on islands off southern Namibia and the West and South Coasts of South Africa, usually from September to November. The birds range widely during the non-breeding season, following their prey (largely sardine and anchovy), which they capture by plunge-diving. The Cape cormorant breeds mostly on nearshore islands and guano platforms, but also at islands within estuaries and lagoons, sewage works and on mainland cliffs. The breeding distribution extends from northern Namibia to Algoa Bay. They are generally dependent on large surface schools of fish, which they capture by pursuit-diving, and do not forage as widely as gannets. Anchovy and sardine are the preferred prey species, with bearded goby an important prey in southern Namibia. The breeding range of African penguins extends from Algoa Bay to Sylvia Hill, south of Walvis Bay. They generally breed on islands, although there are a few small mainland rookeries in South Africa and Namibia. Pelagic shoaling fish, particularly sardine and anchovy, are the most important prey, which are caught by deep pursuit-diving. As with the Cape cormorant, the bearded goby has at times been an important prey off Namibia, particularly when sardine have been scarce. 4. HISTORY AND CURRENT STATUS OF THE FISHERIES Industrial catches of the major species in each country in 1997 are summarised in Table 1. Note that zeroes in parenthesis indicate an assumed zero catch in the absence of data to the contrary. A more general discussion of the history and current status of each sector of the fishery follows.
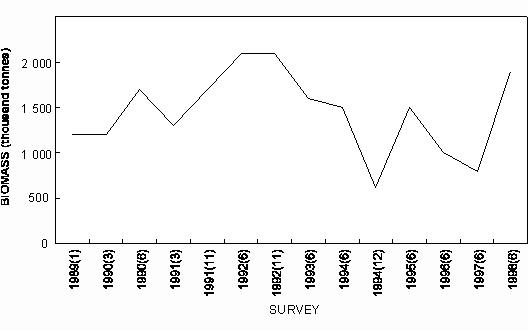
Table 1. Industrial catches (in tonnes) of major species in each country in 1997, except where indicated otherwise. Assumed zero catches in parenthesis. Data for Angola from IIP. Data for Namibia and South Africa from Stuttaford (1998). 4.1 Purse-seine fisheries Annual landings of the major species exploited in the purse seine fisheries of the region from the 1950s to the present are shown in Figs. 6, 7 and 8 for Angola, Namibia and South Africa respectively. They are discussed by species below.
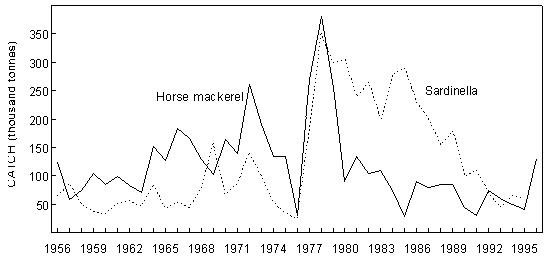
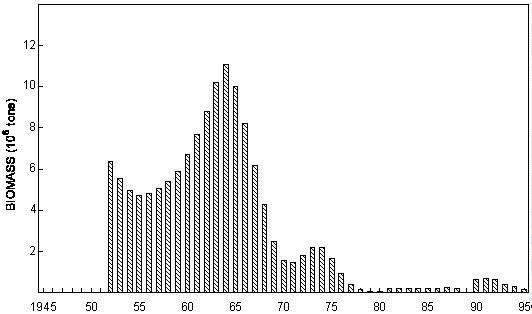
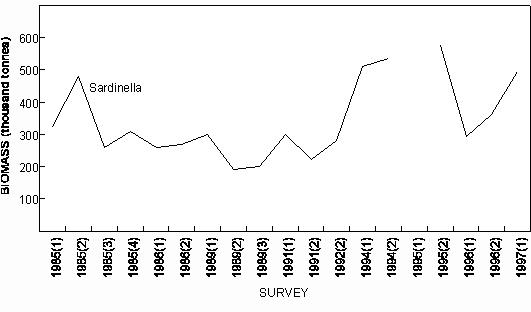
Fig. 6 Purse-seine catches of sardinella (Sardinella maderensis and S. aurita combined) and (predominantly) juvenile Cunene horse mackerel off Angola since 1956
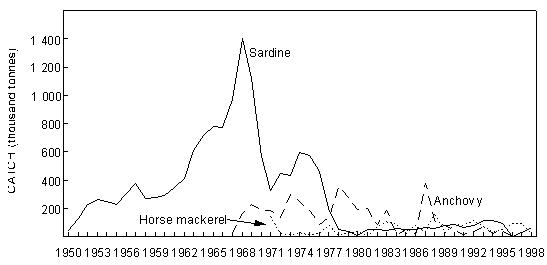
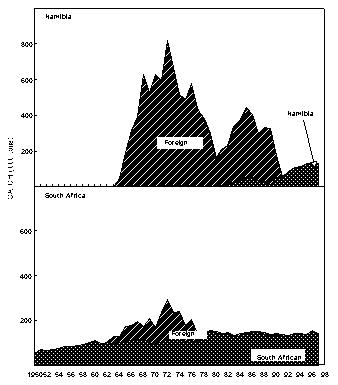
Fig. 7 Purse-seine catches of sardine, anchovy and juvenile Cape horse mackerel off Namibia since 1950
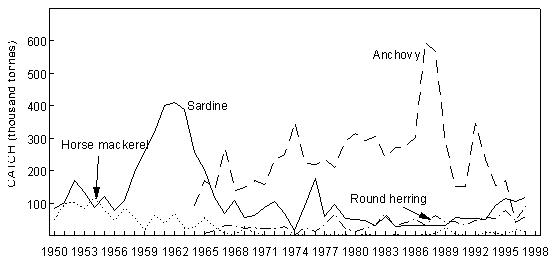
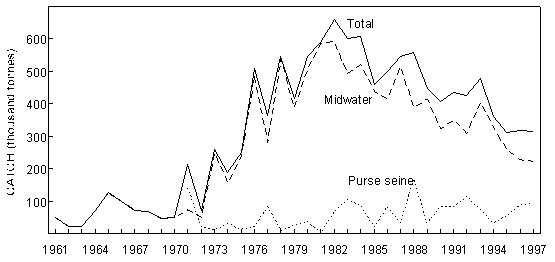
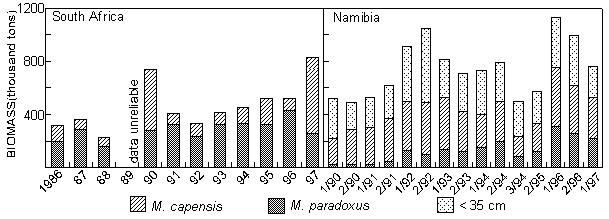
Fig. 8 Purse- seine catches of sardine, anchovy, round herring and Cape horse mackerel off South Africa since 1950 Sardine Industrial catches of sardine in Angolan waters from the 1950s to the present, which are reduced to meal and oil, have fluctuated widely from almost zero to a maximum of 146 000 tonnes in 1957, depending on the state of the Namibian stock and the degree to which it has extended into Angolan waters. The fish were initially caught by a large local fleet of small vessels, but after Independence in 1975 were mainly taken by larger, foreign-owned purse-seiners which began fishing on sardine and sardinellas off Angola in 1976, processing their catches to a large extent at sea. Since 1994, when the adundance of sardine in Namibian waters has been particularly low, a number of Namibian vessels have fished under license for sardine in southern Angola, but only in 1995, when 47 000 tonnes were caught by such vessels, have these catches been significant. In Namibia, annual catches of sardine (mainly adult fish, taken for both canning and reduction to meal and oil) rose rapidly from levels of around 200 000 tonnes to a maximum of nearly 1.4 million tonnes in 1968, whereafter there was a sharp decline to below 300 000 tonnes in 1971, followed by a slight increase in catches for a few years and a precipitous collapse in 1977 and 1978. Since then, annual catches have rarely exceeded 50 000 tonnes, reaching an all-time low of a little over 1 000 tonnes in 1996. It is most likely that these collapses were largely due to over-fishing (especially in the late 1960’s when in addition to the Walvis Bay fleet there were two factory vessels operating outside territorial waters), perhaps exacerbated by a number of years of poor recruitment as a result of adverse environmental conditions. A change from sardine nets to anchovy nets in the late 1960s, which would have placed greater pressure on the recruits, may also have been a contributing factor. With the decline of the stock in the 1970s the fishery moved increasingly northwards, and the fleet changed from small, predominantly wooden-hulled vessels to larger steel-hulled refrigerated-seawater vessels capable of returning the fish from northern Namibia in a condition suitable for canning. South African catches of adult sardine for canning, and of both adults and juveniles for fishmeal and oil, rose from around 100 000 tonnes per annum in the early 1950s to a peak of around 400 000 tonnes per annum in the early 1960s. Thereafter, catches declined sharply to well below 100 000 tonnes per annum, a level which has only recently been regained with the steady growth of the stock since the mid-1980s. Prior to the collapse, the fishery was based on large adults off St Helena Bay, caught predominantly in winter. Thereafter it shifted to younger adults farther south, caught throughout most of the year, and recruits close inshore on the West Coast in autumn and winter as a by-catch in the anchovy fishery, a pattern which still persists. The fleet changed from 32-mm mesh sardine nets to 13 mm anchovy nets in the mid 1960s. Anchovy In the 1970s and 1980s, catches of South African and Namibian anchovy, which were not targeted in South Africa prior to 1964 and were hardly caught in Namibia before 1966, have been less variable than those of sardine, fluctuating under quota control around a level of around 300 000 tonnes in South Africa and 200 000 tonnes in Namibia. An exception was the pronounced peak in 1987 and 1988 in South Africa and in 1987 in Namibia, when catches were roughly double the average. The peak in South Africa is thought to have been due to exceptionally good recruitment, at least partly associated with enhanced transport of eggs and larvae to the West Coast in those years, while in Namibia, the peak was probably caused by an anomalous influx of anchovy recruits from the Cape stock, driven by the same environmental factors which caused the good recruitment in the south that year. Annual catches in South Africa in the 1990s have varied between 40 000 and 347 000 tonnes, but the Namibian catches have averaged less than 50 000 tonnes per annum during this period, with a decline to virtually zero in 1996 and 1997. Recent surveys confirm that the anchovy stock in Namibian waters is severely depleted at present. Horse mackerels The purse-seine fishery for predominantly juvenile T. trecae off Angola started in the 1950s. Catches rose in fluctating fashion from an average of slightly under 100 000 tonnes per annum between 1956 and 1965 to 261 000 tonnes in 1972. Yields then decreased with the cessation of Portuguese rule in Angola, but a record catch of at least 380 000 tonnes was recorded in 1978, by which time distant-water foreign midwater and bottom trawlers, targeting more on adults, had entered the fishery. In the early 1980s, the proportion of the catch taken by purse-seine varied between 15 and 47%, the remainder being taken by midwater (47 - 76%) and bottom (5 - 18%) trawl. Catches declined sharply during this period, and by 1984 only 55 000 tonnes of T. trecae in total were landed. Angolan catches since then have fluctuated between about 25 000 tonnes in 1985 and 1991 and 130 000 tonnes in 1996. A small part of the catch (approximately 2 000 tonnes in 1997 – Anon. 1998a) is taken by small-scale purse-seiners operating close inshore in what is essentially an artisanal fishery. In Namibia, horse mackerel were not recorded in purse-seine landings until 1971 when, following the first collapse of the sardine fishery, 140 000 tonnes were caught. Since then there have been sporadic catches in excess of 100 000 tonnes per year, with an average of 59 000 tonnes and a maximum of 116 000 tonnes in 1992. The fish are utilised entirely for meal and oil. Purse-seine catches of horse mackerel in South Africa, which were once second only to sardine in the landings, peaked at 118 000 tonnes in 1954, and thereafter declined steadily to the early 1970s. Since then horse mackerel have only been a minor constituent of the pelagic fishery, with annual catches never having exceeded 10 000 tonnes. This decline is thought to have been due to a decline in the parent stock, probably aggravated by increased fishing pressure on the younger juveniles with the introduction of anchovy nets in the mid-1960s. Sardinellas Annual catches of the two sardinella species off Angola by the industrial fishery up to the mid-1970s fluctuated between about 50 000 and 150 000 tonnes. With the introduction of the large purse-seiners after Independence, annual catches rose to around 300 000 tonnes, followed by a steady decline from the mid-1980s to 1992 when catches stabilised at around 50 000 tonnes per year. In 1994 there were 36 vessels operating in this fishery, of which 26 were purse-seiners and 10 pelagic trawlers. The catch is used for fishmeal because the fish is too bony for canning. A small amount of sardinella (around 2 000 tonnes in 1997 – Anon. 1998a) is also caught by artisanal fishermen in small-scale purse- seine operations close inshore for local sale and consumption. Round herring Round herring have been a minor constituent of the South African purse-seine fishery since the late 1950s with annual catches in the 1980s and 1990s running at a level of about 50 000 tonnes, peaking at 76 000 tonnes in 1995. The juvenile fish are caught as a by-catch in the fishery for juvenile anchovy and sardine inshore on the West Coast, and the adults in targeted fishing between Cape Columbine and Cape Point, somewhat farther offshore, particularly during the first three months of the year. There have also been sporadic attempts to exploit the large adults in the Algoa Bay region by both purse-seine and midwater trawl. Some of the catch has been canned and the rest reduced to meal. The round herring resource in South African waters is believed to be under-utilised at present, and attempts at greater exploitation have been encouraged. In Namibia, around 1 000 tonnes of juvenile round herring are usually taken each year by the purse-seine fleet for reduction, although catches as high as 14 000 tonnes (in 1996) have been recorded. The potential for canning the larger fish has been investigated, but they have generally been found to be too soft for this purpose. 4.2 Trawl fisheries Hakes Annual catches of Cape hakes (M. capensis and M. paradoxus combined) in Namibian and South African waters by local and foreign fleets since 1950 are shown in Fig. 9. Although the demersal fishery began around the turn of the century, catches prior to 1950 seldom exceeded 50 000 tonnes per annum, with most fishing effort being in South African waters. The Namibian fishery started in the late 1950s. In the early 1960s there was an explosive increase in effort and landings throughout the Benguela, with the arrival of foreign trawling fleets, and by 1972, the annual hake catch in the south-east Atlantic exceeded 1.1 million tonnes. Subsequently, catch rates and landings of hake declined sharply, and conservation measures were introduced, including the declaration of a 200-mile fishing zone by South Africa in 1977. Since then hake catches in South African waters have remained relatively stable at just over 140 000 tonnes per year. Off Namibia, hake catches between 1973 and Independence in 1990 averaged 500 – 600 000 tonnes annually, mainly taken by foreign fleets. At Independence, strict conservation measures were introduced, including the exclusion of foreign vessels. The hake catch is now taken exclusively by Namibian-registered vessels and the annual local catch has risen from 55 000 tonnes at Independence to around 120 000 tonnes over the period 1996 – 1998. Catches of M. capensis and M. polli in Angola are of a lower order, amounting to less than 1 000 tons per year in recent years.
Fig. 9 Catches of hakes off Namibia and South Africa by foreign and local fleets since 1950 Horse mackerels Adult horse mackerel are the main target for midwater trawlers operating in Namibia and Angola, the Namibian fishery being the largest by volume in that country. Trawl catches of the two species rose from under 50 000 tonnes per annum in the early 1960s (when horse mackerel were only trawled in Namibia), to between 600 000 and 700 000 tonnes per annum from 1982 to 1984 (Fig. 10), by which time most of the catch was being taken by foreign (mainly former Soviet bloc) vessels. The fish were largely frozen and shipped back to Europe for human consumption. Trawl catches of horse mackerel (mainly T. trachurus capensis) in Namibia since Independence in 1990, when Namibia took control of the fishery, have fluctuated around 350 000 tonnes per annum, with a decline to between 200 000 and 250 000 tonnes per annum in recent years. The number of midwater trawlers in the fishery now is less than half that at Independence. The fleet is largely made up of ageing ex-Soviet bloc vessels, about half of which are now registered in Namibia, but which are mostly operated by foreign crew. The major part of the catch is frozen and transhipped to reefer vessels for export as a relatively low-value product to West Africa, but a small amount is now being smoked or dried-salted ashore for export to African countries.
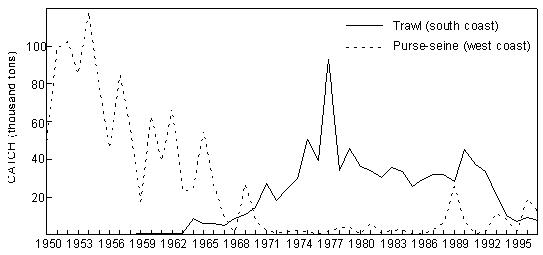
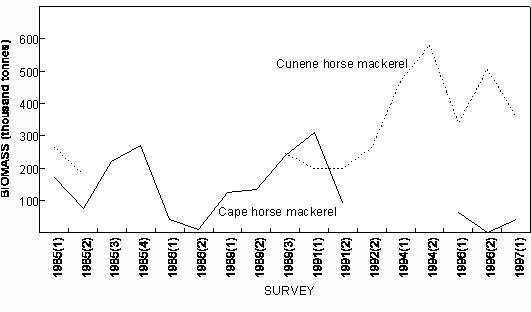
Fig. 10 Catches of (predominantly) Cape horse mackerel by midwater trawl and purse-seine off Namibia since 1961 Purse-seine and trawl catches of Cunene horse mackerel in Angola were summarised in Section 4.1. Compared to these catches, which have generally been between 50 000 and 100 00 tonnes per annum over the past decade, catches of Cape horse mackerel in Angolan waters in the past decade have been insignificant. The fishery for T. trachurus capensis in South Africa changed from a purse-seine operation for juveniles on the West Coast in the 1950s and early 1960s to a bottom-trawl fishery for adults on the South Coast thereafter, caught by both local and foreign (mainly Japanese) trawlers. Catches rose from about 10 000 tonnes in 1964 to a peak of nearly 100 000 tonnes in 1978 (Fig.11), and have averaged about 30 000 tonnes per annum over the past two decades. Between 1967 and 1975, horse mackerel contributed some 40% of the landings of the inshore demersal fishery on the South Coast, but since the gradual phasing out of the foreign fishery from 1982, this proportion has declined to around 20%. In the 1990s a targeted midwater trawl fishery for horse mackerel developed, mostly on the Eastern Agulhas Bank. Catches have been relatively low (average just under 8 000 tonnes per annum between 1990 and 1997) due to operational and marketing constraints, but there is concern that this fishery has the potential to threaten the resource. Horse mackerel catches in bottom trawls on the West Coast in the past decade have never exceeded 5 000 tonnes per year.
Fig. 11 Cape horse mackerel catches in the South African purse-seine (West coast) and trawl (South Coast) fisheries since 1950 Monkfish The Namibian monkfish fishery was initially a by-catch fishery, but in recent years it has developed into a target fishery in response to increasing market demand. At Independence in 1990 the fishery changed from an international to a local fishery. Since then annual catches (mainly by small wetfish trawlers) have risen from 1 500 tonnes (the lowest value in the past decade) to around 10 000 tonnes, comparable to peak levels reached by the international fishery in the 1980s. The value of the product is high, making this fishery an important contributor to the Namibian economy (Olsen 1997). The by-catch of monkfish in the South African demersal trawl fishery on the West and South Coasts has been relatively constant over the past two decades, at around 5 000 tonnes per year. Other trawled species In Angola, roughly 10 000 tons of demersal fish have been caught annually in trawls in recent years, mainly dentex spp., croakers (Scianidae), grunters (Pomadasydiae) and groupers (Serranidae). In Namibia the most important other species caught in bottom trawls are kingklip and sole, catches of which have averaged 1 275 and 340 tonnes respectively per year since 1990. The average combined catch of dentex and jacopever in midwater trawls in Namibia since 1990 is 3 750 tonnes (Boyer et al. 1998). In South Africa the two most important other trawled species are snoek and kingklip. Between 1990 and 1995 catches of these two species averaged 10 900 and 2 200 tonnes per year respectively. Deep-water species Exploratory fishing for deep-water trawl species, particularly orange roughy, began in Namibia in 1994. Small catches were made in 1995, but in the following season over 12 000 tonnes of roughy were caught (by bottom trawl) making this the second-largest roughy fishery in the world. The alfonsino catch during these two years was nearly 3 000 tonnes. Indications are that the 1997/98 catch of both species will be at the same level. At present only four roughy aggregations are being targeted, although there is some exploratory fishing for further aggregations. Five vessels are currently operating, in joint ventures between foreign and Namibian companies. The roughy and alfonsino catch is entirely exported, mainly in the form of high-value frozen fillets for the USA and Japanese markets respectively. 4.3 Crustacean fisheries West Coast rock lobster There has been a fishery for West Coast rock lobster since the early part of the century, and for a long period the fishery was the world’s largest for any Jasus species. During the 1940s and 1950s South African catches were relatively stable at around 9 000 tonnes (whole mass) per annum, but from the mid 1960s to the present catch levels have declined under quota control to around 1 500 tonnes per annum, with particularly sharp declines in the late 1960s, early 1980s and early 1990s, the most recent one being attributed at least in part to a sharp decline in the growth rate of individuals. In Namibia, where the resource was clearly over-exploited, annual catches have declined even more dramatically, from a peak of nearly 9 000 tonnes in 1966 to about 3 000 tonnes in the early 1970s, to half this level a decade later, and to a few hundred tonnes in recent years. Deep-sea red crab The red crab fishery in Namibia started in 1973. Catches rose to a peak of about 10 000 tonnes in 1983, after which annual landings declined steadily to 2 676 tonnes in 1991, clearly as a result of over-fishing. Catches have fluctuated around this level since then. In only two of the years since the introduction of TACs in 1989 has the TAC been reached. The entire catch is taken by Japanese vessels fishing with traps. Off Angola, a few hundred tonnes of red crab are taken by trawl each year as a by-catch of the deep-water prawn fishery. Since 1986 there has also been a directed crab fishery by a single Japanese vessel, which has caught a few thousand tonnes per year on average. Deep-water Prawns The deep-water prawn fishery off the west coast of southern Africa (mainly for rose and striped prawns) began in 1966, when a single Spanish trawler started operating off Angola. By 1972, a fleet of 54 foreign (mainly Spanish) trawlers was operating off Angola, and catches had risen from about 1 000 tonnes per annum to over 8 000 tonnes per annum. The catch peaked at over 12 000 tonnes in 1973, whereafter there was a sharp reduction in Spanish operations, ending in complete withdrawal from the fishery by 1977. The Spanish fleet was replaced by a Cuban fleet, which continued to fish until 1979, the total catch peaking at 11 400 tonnes in 1975. Since then catches have declined, with annual landings of the two species combined over the past decade fluctuating between about 3 500 and 7 000 tonnes per year (Fig. 12) peaking in 1997. In all but two of the years, landings of rose prawn have been higher than those of the striped prawn. At present, there are 44 vessels in the fishery, of which 22 are fishing under an agreement with the European Union, and the remainder are national.
Fig. 12 Annual landings of deep-water rose prawn and striped prawn in Angola since 1988 (from Sardinha) 1998) 4.4 Commercial linefisheries The total annual catch of tuna in Angolan waters between 1993 and 1995 reported to ICCAT varied between 291 and 910 tonnes, with yellowfin tuna (caught by local baitboats and longliners) and big-eye tuna (caught by foreign longliners) making up most of the catch . Foreign longliners caught tuna in Namibian waters under South African licence prior to Independence in 1990. The Namibian-controlled tuna fishery started in 1991, and since then an average of 2 330 tonnes of tuna (predominantly southern albacore) have been taken per year by a fleet of about 30 local and foreign-owned pole or line vessels. The foreign longline tuna fishery, which targets bigeye tuna for the high-value sashimi market, started in 1993. Catches have varied between 52 tonnes in 1996 and 1 005 tonnes in 1994. Experimental catches of swordfish Xiphias gladius taken by surface longlining have been low (50 tonnes in three years), but are regarded as encouraging given the low level of effort. An average of 770 tonnes of snoek has been taken per year by commercial vessels since Independence, mostly by handline (Boyer et al. 1998). Catches of silver kob from commercial linefish vessels have fluctuated over the past three decades around an average of about 500 tonnes per year. It is estimated that at present the recreational catch of silver kob is roughly equal to the commercial catch (Kirchner 1998). The commercial linefishery in South Africa developed rapidly after the Second World War, reaching its peak in the late 1960s, when about 20 000 tonnes on average were caught per year. Thereafter, catches declined despite increasing effort. The average annual catch of all species excluding tuna between 1993 and 1997 was 11 700 tonnes, of which just over half was caught west of Cape Point. Snoek have consistently contributed over half the catch since the start of the fishery. The average catch over the past five years was 6 650 tonnes, of which more than half was landed at St Helena Bay and Yzerfontein on the West Coast. The second-most important species in terms of landings was hake, with average landings of just under 1 000 tonnes per annum, caught mostly on the South Coast. Other important species include yellowtail Seriola lalandi, kob spp. (particularly Argyrosomus inordus) and geelbek Atractoscion aequidens, which together contribute almost as much as snoek to the annual landings. There is a large and varied recreational fishery for linefish species in South African waters, particularly on the South and East Coasts. Off the West Coast, the species most commonly taken from boats are tuna, snoek and yellowtail, while galjoen, hottentot, silver kob and white stumpnose Rhabdosargus globiceps are the bony fish most commonly caught by shore-anglers. The baitboat (pole) fishery for tuna in South African waters started in 1980, and by 1990 about 10 000 tonnes of tuna (predominantly southern albacore) were being caught per year. Catches subsequently declined to about half this (6 571 tonnes on average between 1993 and 1997), but reached about 8 000 tonnes again in 1998. For the past two decades there has also been a longline fishery for large bigeye and yellowfin tuna by Japanese and Taiwanese vessels fishing under license, catches of the latter species being confined to the warmer waters east of Cape Point. In 1997, experimental pelagic longline permits were issued to South African fishermen, primarily to target high-grade tuna for fresh export. The vessels have made good tuna catches along the edge of the continental shelf, and have also caught substantial quantities of swordfish (probably nearly 1000 tonnes in 1998), which has a high market value and is likely to be exploited more heavily in the next few years. 4.5 Artisanal/subsistence fisheries By far the most important artisanal fisheries in the region are in Angola, where they contribute some 30% of the total landings of fish and shellfish. Declared landings from this sector in 1997 amounted to nearly 30 000 tonnes (Anon. 1998a). In 1995 there were more than 23 000 registered artisanal fishermen, landing their catches at some 105 controlled landing places, predominantly in the central provinces of Luanda, Benguela and Kwanza Sul (Delgado and Kingombo 1998). The total number of boats in 1995 was estimated at 4 677, ranging in size and sophistication from unmotorised canoes (about 25% of the total) to small wooden boats 5 - 6.5 m long with or without motors ("chatas"), which make up about 70% of the total, to 8 – 12 m vessels ("catrongas") with inboard motors and some preservation facilities (Delgado and Kingombo 1998). Larger "traineiras" (8 – 25m semi-industrial deckboats with inboard engines) are also involved in small-scale fisheries along the coast. The most common fishing gears are gillnets, longlines, and beach- and boat-operated seine nets and traps. The main species caught are pelagic fish such as sardine and sardinellas, horse mackerel, mullet, bigeye grunter and small tuna, and demersal fish such as bigeye dentex, croakers and groupers. Fresh fish is sold in urban areas close to the landing sites, or because of a lack of freezing facilities, is dried or smoked for sale in the rural areas. Artisanal/subsistence fisheries off the West Coast of South Africa (particularly for mullet Liza richardsoni, which is now caught mainly by beach-seine and driftnets), have been in existence for centuries, but are of negligible value compared to the nation’s major industrial fisheries. In Namibia there is virtually no artisanal/subsistence fishing, because of the uninhabited desert coastline. 4.6 Recreational fisheries Marine recreational fishing from skiboats and larger gamefishing boats, and by rock-and-surf anglers and spearfishermen is an important leisure activity in South Africa, and directly or indirectly generates a significant amount of employment and revenue (see Section 7). The sector has grown rapidly since the Second World War, to the extent that surveys indicate that approximately 15% of coastal residents now fish in the sea on a regular basis. As the fishery has grown, there has been a tendency along the entire coast for catch rates to decline, and for endemic inshore reef species (such as galjoen and white stumpnose in the Western Cape) to be replaced by cartilaginous fish in the rock-and-surf catches. Recreational fishing in Namibia is pursued by rock-and-surf anglers, who have access to about 20% of the coastline, and skiboat fishermen operating from Walvis Bay, Swakopmund and Lüderitz. The sector has grown rapidly in recent years, and now attracts many visitors to the coast, particularly from Namibia and South Africa, generating considerable employment and revenue in the coastal towns (see Section 7). With the recent increase in fishing pressure, the number of fish caught per angler has declined significantly. 5. CHANGES IN ABUNDANCE AND DISTRIBUTION OF MAJOR RESOURCES 5.1 Pelagic resources Sardine and anchovy Virtual Population Analysis (VPA) estimates of sardine biomass in Namibia between 1952 and 1988 (Fig. 13) confirm the marked decline of the stock in the second half of the 1960s, and the collapse to well below 1 million tonnes in the mid 1970s. Catch patterns have clearly indicated a northward shift in the core distribution since the collapse of the fishery in the 1970s, possibly as a result of the depletion of the southern spawning population and the cessation of associated migrations. Acoustic survey estimates of sardine on the Namibian/southern Angolan shelf since 1990 (Fig. 14) indicate that the adult stock is still well below pre-collapse levels, with an all-time lowest estimate of only a few thousand tonnes in the summer of 1995/1996, co-incident with the Benguela Nińo at that time (Section 6.1). The surveys also indicated a further northward shift in distribution in 1994, when the majority of the biomass was found off southern Angola. Survey estimates of anchovy and juvenile round herring combined (they are not distinguished in the surveys) have dropped from levels of around 200 000 tonnes between 1990 and 1993 to below 100 000 tonnes since then. The decline is reflected in the anchovy landings, which have been negligible since 1996.
Fig. 13 VPA estimates of sardine biomass in Namibian waters from 1952 to 1988 (from Boyer 1996)
Fig. 14 Estimates of sardine biomass in Namibia/southern Angola since 1990, obtained from acoustic surveys (updated from Boyer 1996). * = Angola not surveyed. In South Africa, acoustic surveys of sardine over the past 14 years have revealed a gradual increase in spawner biomass from below 50 000 tonnes in 1984 to around 600 000 tonnes at present (Fig. 15). The same surveys (supported by concomitant egg-production surveys between 1983 and 1990) have revealed two major peaks in anchovy spawner biomass in South African waters during this period, with pronounced troughs in 1989/1990 and in 1996 (Fig. 15). Acoustic surveys of sardine and anchovy recruitment off South Africa show the same general pattern (Fig. 16), except in 1995, when strong anchovy recruitment was not matched by a high spawning biomass in that or the previous year. Of particular note is the high degree of correlation between the sardine and anchovy recruitment estimates throughout the time-series, which suggests that, in South Africa at least, recruitment of these two species is controlled largely by common environmental influences on the early life history stages, and not primarily by spawning biomass.
Fig. 15 Acoustic estimates of anchovy and sardine spawner biomass off South Africa since 1984 (from Barange et al. in press) The South African surveys have also revealed significant shifts in sardine and anchovy distribution in the Southern Benguela since the early 1980s, the most prominent of which was a sudden large increase in the number of anchovy spawners on the West Coast in 1986, followed by a progressive reduction to the more normal low levels over the next three years (Hampton 1992). A significant eastward shift in both the anchovy and the sardine spawning populations has also been recorded in the past decade, from 1990 in the case of anchovy, and from 1988 onwards for sardine (Barange et al. in press).
Fig. 16 Acoustic estimates of the number and biomass of anchovy and sardine recruits off South Africa since 1995 (from Barange et al. in press) Sardinella Dr Fridtjof Nansen acoustic survey estimates of the biomass of sardinella (S. aurita and S. maderensis combined) in Angolan waters between 1984 and 1997 are shown in Fig. 17, which suggests that sardinella biomass has increased from levels of around 200 000 tonnes in the 1980s to more than double this in the 1990s. (This increase was probably at least partly due to the withdrawal of a major part of the distant-water foreign fleet in the late 1980s). It should be noted that at least some of the survey estimates are possibly negatively biased because of the difficulty in assessing sardinella acoustically when the fish are on or close to the surface, which frequently occurs.
Fig. 17 Estimates of sardinella (S. maderensis and S. aurita) biomass in Angola since 1985, from Dr Fridtjof Nansen acoustic surveys (Data from IIP) Round herring According to acoustic survey data, round herring spawner biomass in South Africa appears to have remained relatively constant at around 1 million tonnes since the first estimate was made in 1986. No pronounced shifts in the distribution of adults have been noted from the survey data, although since the surveys have not always encompassed the distributional range of the population, the possibility of such shifts cannot be excluded. 5.2 Trawled species Hakes Survey estimates of M. capensis and M. paradoxus abundance off South Africa and Namibia over the past decade, derived from swept-area demersal surveys (supplemented by acoustic estimates of fish above the trawl in the Namibian surveys), are shown in Fig. 18. It is notable that there has been a marked increase in the abundance of M. paradoxus off Namibia since 1992, which is confirmed by a marked increase in the proportion of M. paradoxus in the Namibian hake catches in recent years (E. Voges, NatMIRC, pers. comm.). This increase may indicate northward displacement or expansion of the stock from South Africa, or alternatively, a shoreward displacement in response to changes in the oxygen content of bottom waters. Fig. 16 indicates that in Namibia, the component of the population targeted by the fishery (i.e. M. capensis >35 cm long) grew between Independence in 1990 and 1992, but thereafter declined for the next four years. The most recent survey results indicate exceptionally good recruitment in 1997 (Boyer et al. 1998), with a subsequent increase in fishable biomass to the highest level since Independence. Off South Africa, the survey results suggest an increase in M. paradoxus biomass on the West Coast over the past decade, but no clear trend in the biomass of M. capensis. The low estimate for M. paradoxus in 1992 supports the hypothesis of a longshore displacement of the Cape stock into Namibian waters in that year, but given the uncertainty in the estimates, this apparent connection should be viewed with caution.
Fig. 18 Research survey estimates of the biomass of Cape hakes (Merluccius capensis and M. paradoxus) off the West Coast of South Africa and off Namibia (data from M&CM and NatMIRC respectively) Fig. 19 (from Anon. 1998b) shows trends in exploitable and total biomass of hake on the West Coast of South Africa from 1917 to the present, estimated from an age-structured production model. All indications are that the stock has been relatively stable over the past two decades, with signs of a gradual increase in recent years.
Fig. 19 Estimates of the exploitable and total biomass of Cape hakes in South African waters since 1917 from an age-structured production model Horse mackerel Acoustic estimates of horse mackerel (mainly T. trecae) biomass in Angolan waters between 1985 and 1997, obtained from Dr Fridtjof Nansen surveys, are shown in Fig. 20. There appear to be two distinct periods: from 1985 to about 1995, when T. trecae biomass fluctuated around a level of about 200 000 tonnes and capensis biomass was occasionally of the same order, and from 1995 to the present, when T. trecae levels have been about twice as high, and T. t. capensis biomass very low. In all surveys the abundance was highest in central and southern Angola, except in 1996 (a year of anomalously warm conditions along the whole Angolan coast) when a third of the biomass was found in the north, between Luanda and Cabinda. The level of these estimates compared to the catches made at the height of the Angolan horse mackerel fishery (over 380 000 tonnes in 1978) is clear indication of the decline of the resource, which is supported by the more than 10-fold reduction in annual catches which has occurred since then (Section 4.1).
Fig. 20 Acoustic survey estimates of Cunene and Cape horse mackerel biomass in Angolan waters since 1985. Data from IIP (NB: check this Fig!) Using a VPA of catch data from the former Soviet bloc fleet, Vaske et al. (1989) estimated that the biomass of Trachurus spp. in ICSEAF Divisions 1.3, 1.4 and 1.5 (i.e. between southern Angola and Cape Agulhas) between 1973 and 1987 fluctuated between about 1.5 and 2.3 million tonnes. This estimate must however be treated with caution because of, inter alia, the unreliability of the age-length keys used in ageing the fish. Since 1990, the biomass of adults and juveniles combined off Namibia and southern Angola has been estimated by annual acoustic surveys, predominantly from Dr Fridtjof Nansen (Fig. 21). The estimates generally fall between 1 and 2 million tonnes, with a maximum of 2.1 million tonnes in 1992. In an absolute sense, these estimates are questionable because of the lack of verification of the target strength expression used. For example, use of a target strength expression for T. trachurus capensis developed in South Africa (Barange and Hampton 1994) would lower the estimates by a factor of approximately 3. Furthermore, some of the differences between the surveys have been attributed to variations in the proportion of the population which is acoustically detectable rather than to fluctuations in the biomass (Anon. 1998c) – a problem which has also been experienced in acoustic surveys of horse mackerel in South Africa (see below).
Fig. 21 Acoustic survey estimates of (predominantly) Cape horse mackerel biomass in Namibian waters since 1989. Data from NatMIRC Bottom trawl survey estimates of T. trachurus capensis biomass on the South African West Coast between 1985 and the present have increased from less than 10 000 tonnes in 1986 to between 50 000 and 300 000 tonnes since 1991. Equivalent estimates on the South Coast over this period for the 0-200m depth stratum have fluctuated between 100 000 and 580 000 tonnes, with an average of about 300 000 tonnes. There is however evidence from acoustic surveys and other acoustic observations that the South Coast trawl estimates are negatively biased because of the unavailability of part of the population to the bottom trawl at certain times and in certain areas (particularly on the outer shelf and over the shelf-break). Deep-water fish From a swept area analysis of commercial catches in 1996 and 1997, Branch (1996) and Branch and Roberts (1998) estimated the total abundance of orange roughy on the Namibian shelf at that time at over 200 000 tonnes. No biomass estimates considered reliable have yet been obtained for alfonsino. An acoustic survey of the three most important roughy grounds from Dr Fridtjof Nansen in 1997 gave an estimate of 147 000 tonnes after correction for the major sources of bias. A major difficulty in assessing the total roughy population by either trawl-based or acoustic methods is the lack of knowledge regarding the proportion of the population outside the known aggregations. 5.3 Crustaceans West coast rock lobster The Schaefer and Fox production models used to estimate the MSY for the Namibian rock lobster resource (Section 8.4) have given values of 4 300 and 3 100 tonnes respectively for the period 1958 - 1991. If the very variable data from the 1960s and the TAC-restricted data from the early 1990s are ignored, these estimates drop to 1 900 and 1 800 tonnes respectively. Recent modeling estimates put the stock at around 3 000 tonnes at present. Assessments of the South African rock lobster resource based on conventional size-based analyses have shown it to be seriously depleted, estimates of recruitment in recent decades being only some 35% of pristine. Deep-sea red crab Photographic surveys of red crab within two approximately 50-km long sections of the northern Namibian shelf in the early 1980s indicated a biomass of approximately 5 000 tonnes in the one area and about 2 000 tonnes in the other. Recent estimates from analytical models (Section 8.4) indicate that the Namibian component of the stock has declined from about 40 000 tonnes at that time to around 10 000 tonnes in the 1990s, which is reflected in the decline in the catch rate during this period (Section 4.3). In Angola, estimates of 47 6000 tonnes, 91 000 tonnes and 18 000 tonnes have been reported for 1977, 1982 and 1996 respectively (Neto 1997), perhaps indicating a decline of the Angolan stock as well, which might be expected in view of the fact that this appears to be a single shared stock, with considerable trans-boundary migrations. Deep-sea prawns Estimates of rose prawn and striped prawn abundance off Angola have been made from Dr Fridtjof Nansen trawl surveys between 1985 and 1997. The estimates for the two species combined vary between 1 770 tonnes in 1996 and 5 850 tonnes in 1986, divided almost equally between the two species. There does not appear to have been any major shift in distribution during the survey period. 5.4 Line-caught species Judging from commercial handline catches, the abundance of snoek in Namibia increased some three-fold between 1970 and 1980, probably in response to an increase in the abundance of prey in the form of juvenile horse mackerel, which was then the dominant pelagic fish species in the region. This increase was also reflected in catches of snoek by the international midwater trawl fleet operating off Namibia at the time, which rose sharply during this period to levels of over 20 000 tonnes per year. Relatively constant catches by commercial linefishermen and in trawls since Independence suggest that the population is relatively stable at present, although the trawl catches are now an order of magnitude lower than in the 1970s and 80s. It is not clear whether the latter reflects a major reduction in biomass, or is due to a change in distribution and/or fishing strategy. Commercial catches of other linefish species in Namibia have also not fluctuated widely since Independence. Kirchner (1998) has recently estimated that the current exploitable biomass of the most important of these (silver kob) is in the region of 11 000 tonnes. The estimate was obtained from a Thompson and Bell yield-per-recruit model, using catch-at-age data for two years. She estimates that the stock is currently at between 29 and 46 % of its virgin biomass and that the current fishing mortality is between 0.12 and 0.22 yr-1. Fluctuations in the abundance of line-caught species in South Africa are evident from catch-per-unit-effort (CPUE) indices, which for practically all species have declined over the past few decades, to almost zero in some cases. This is the case for both resident reef fish and migratory shoaling species. For many of the latter it has been estimated that spawner biomass/recruit ratios are now below the recommended critical threshold level of 25% (Penney et al. 1997), with ratios for important species such as the silver kob being only 3 – 12% of pristine. As a result of stock declines, the distributional ranges of many of these species appear to have contracted towards the centre of their past distributions, and the magnitude and extent of their migrations has declined. Of the migratory shoaling species, snoek and yellowtail are perhaps the only ones considered not to be overexploited in South African waters, although there have been substantial fluctuations in abundance and distribution of both species in the past, probably in response to changes in the abundance and distribution of their pelagic prey. 5.5 Seals and seabirds The Southern African seal population was heavily overexploited in past centuries and the breeding colonies around the Cape Peninsula were exterminated soon after the arrival of European settlers in the 17th century. Subsequently, as a result of conservation measures applied over many years, and probably the establishment of new mainland colonies in areas of restricted human access, the population increased from around 100 000 animals in 1900 to 1.5 to 2 million animals in the early 1990s, as deduced from aerial surveys of pups and tag-recapture studies. The Namibian population subsequently declined (probably by almost a factor of 2) as a result of breeding failure and high pup and adult mortalities brought about by the effects of the 1994-95 intrusion of warm, low-oxygen water into northern Namibia (Anon. 1997a). In 1994 and 1995 there was also a northward shift in distribution, with Cape fur seals being found as far north as Luanda, feeding on sardinella. Recent information suggests that the distribution has now returned to normal, and that the population has almost recovered to previous levels (Geromont et al. in press). Aerial censuses have shown that between 1956 and 1986, the population of Cape gannets at the Namibian colonies declined from around 200 000 adults to only about 50 000, which was probably due to the decline and subsequent collapse of the Namibian sardine resource during this period. Over the same period the adult populations at the two colonies off the Westen Cape remained relatively constant at around 30 000 birds, whereas numbers at Bird island, Algoa Bay, increased from around 20 000 to approximately 60 000 birds. The surveys also revealed a substantial decline in the numbers of Cape cormorant, from more than a million birds in the early 1970s to about 277 000 pairs in the late 1970s, and only about 120 000 pairs in the mid 1980s. The decline was most severe in the north. As with the gannet, the population on the South Coast increased as the West Coast population declined, by a factor of five between 1956 and 1978. The African penguin population in Southern Africa has been reduced to very low levels during the last few centuries through prolonged and excessive egg collection and disruption of their breeding habitats during the guano rush in the 1840s. The most recent estimate of the total population, from an aerial census in 1995, is about 150 000 birds, of which about 40% were found on islands off the South African West Coast. Even allowing for underestimation owing to inadequacies of the aerial survey technique, this population is probably at least an order of magnitude below pre-exploitation levels. As with Cape gannets and Cape cormorants, the distribution of penguins has changed over the past three decades in response to changes in the distribution and relative abundance of sardine and anchovy (Crawford 1998). Crawford and Dyer (1995) and Crawford (in press) have shown that the proportion of sardine and anchovy in the diet of seabird in the southern Benguela is strongly correlated with acoustic estimates of sardine and anchovy abundance. An example is shown in Fig. 22, which compares the trends in the diets of gannets with the acoustic estimates of sardine and anchovy spawner biomass since the start of the surveys in 1984.
Fig. 22 Acoustically derived estimates of sardine and anchovy spawner biomass off South Afica since 1984, compared with the contribution of these species to the diet of gannets in the region (from Crawford in press) 6. EFFECTS OF THE ENVIRONMENT ON DISTRIBUTION AND ABUNDANCE Although a number of Benguela resources have clearly been over-exploited in the past, some of the variations in abundance and distribution which have been observed are more likely to have been caused by the major perturbations which have occured in both the northern and the southern Benguela over the past three decades, and which are summarised in Thematic Report 2. The probable effects of these perturbations on some of the major resources of the region are discussed briefly below. 6.1 Pelagic resources In a recent Principal Components Analysis of satellite SST imagery from the northern Benguela between 1981 and 1987, Cole and McGlade (1998) identified three spatial/temporal patterns which characterised the physical dynamics of the system. They related two of them (the balance between cross-shelf and longshore SST gradients, and the warming of the central region in relation to conditions to the north and/or south), to conditions favourable for clupeoid production in the region. Other, more specific impacts of the environment on pelagic resources of the region have been observed, as detailed below. The biomass of sardine in both the northern and the southern Benguela declined sharply following a system-wide Benguela Nińo in 1963, which in the northern Benguela caused the fish to be concentrated close to Walvis Bay where fishing pressure was high. The subsequent collapse in the Namibian sardine after 1974 (Fig. 14) followed a protracted but less intense local Benguela Nińo between 1972 and 1974, whose effects were probably aggravated by over-fishing. O’Toole and Shannon (1997) have postulated that the recent decline in Namibian sardine, which started in 1993, was largely the result of the advection of low-oxygen water from Angola in 1993 and 1994, aggravated by a major Benguela Nińo in 1995, which also originated off Angola. In March of that year, the Benguela Nińo caused the entire coast from Cabinda to central Namibia to be covered by anomalously warm water (up to 8 oC above average in places) to a distance of more than 300 km offshore (Gammelsrřd et al. 1998). Observed mortalities of sardine, horse mackerel and silver kob, and poor recruitment and declining catch rates of a number of other key resources in Namibia at the time, are further indications of a broad-scale environmental effect on sardine and other resources in Namibia at the time. There was also increased fishing pressure on sardine, caused by a southward displacement of sardine from northern Namibia and Angola, increasing the availability of these fish to the Walvis Bay fleet. The more gradual increases in sardine abundance in the southern Benguela since the mid-1980s, and to a lesser extent in the northern Benguela in the late 1980s, may be related to the fact that this was a relatively warm period, with no major environmental perturbations. Although these observations are all highly speculative, they are suggestive of strong relationships between sardine biomass and the environment in both the northern and the southern Benguela which bear further investigation. In the case of anchovy, a Benguela Nińo in 1984, which followed an extended cold period on the shelf, seems to have had an adverse effect on the Namibian resource. As suggested in Section 3.1, the exceptionally good catches in 1987 were probably largely the result of recruitment from the strong year-classes of 1986 and 1987 in the southern Benguela, rather than from a recovery of the Namibian stock. Following the most recent Benguela Nińo in 1995, which came at the end of a period of low abundance (Fig. 7), the Namibian anchovy resource appears to have collapsed completely, or perhaps to have been displaced. In the southern Benguela, the exceptionally good anchovy recruitment in 1986 and 1987 is thought to have been at least partly associated with enhanced leakage of Agulhas Bank shelf water onto the West Coast, and the increased influence of westerly winds, which would have resulted in favourable transport and entrainment of eggs and larvae with minimum offshore advective loss. The weak anchovy recruitment in 1989 was associated with poor feeding conditions during the latter part of 1988 in the spawning area over the western Agulhas Bank, and with less favourable transport to the nursery grounds on the West Coast. In general, it appears from extensive studies of transport mechanisms, food availability, wind stress, primary production and other environmental influences, that in the southern Benguela there is an optimum environmental window for good anchovy (and probably sardine) recruitment, with too much or too little wind stress, primary production etc. being detrimental to recruitment (e.g. Hutchings et al. 1998). Variations in these conditions may explain much of the observed variation in recruitment (Fig. 16), and hence in spawning biomass, although there is evidence that other factors such as temperature and feeding conditions on the spawning ground, the location and duration of the spawning, and the condition of spawners can also have an effect on recruitment (e.g. Hutchings et al. 1998, Painting and Korrűbel 1998, Korrűbel et al. 1998, Shannon 1998). The balance of the evidence does however suggest that offshore advective loss of ichthyoplankton, which is present to some extent in all years, is probably a major controlling factor in both anchovy and sardine recruitment. Isolating and ultimately quantifying the most important environmental factors affecting the recruitment of pelagic fish in the Benguela region is a major research thrust, both in national research programmes and in regional/international programmes such as BENEFIT, ENVIFISH and VIBES (see Section 8.3). On a larger scale, the meridional distribution of sardine and anchovy in the Benguela Current may be affected by shifts in the major wind belts across the African continent. Their distribution might therefore be expected to be connected to that of sardine and anchovy in the Canary Current, which would also be affected by such shifts. In contrast, there is some evidence that regime shifts in the Benguela, and switches between sardine and anchovy dominance (which according to scale-deposit studies have a characteristic periodicity of around 50 years), tend to be out of phase with those in the Pacific. It is also notable from a comparison between Figs 7 and 8 that, over the past two decades, there has not been a close correspondence between abundance trends in the northern and southern Benguela for either sardine or anchovy. This could possibly be due to the major differences in the geographical relation between the spawning and upwelling areas in the northern and southern Benguela noted in Section 3.1, and consequent differences in the effect of wind stress on recruitment processes. 6.2 Trawled species Relatively little is known about the behaviour of Cape hakes in the Benguela ecosystem, and of their responses to environmental variability and change. Adult hake are good swimmers, undergo vertical migrations, can tolerate a range of temperatures, and are particularly well-adapted to low oxygen conditions, the adults being able to tolerate levels as low as 0.25 ml l-1 . They are therefore well able to react to unfavourable environments, and being opportunistic feeders, long-lived and inhabiting a wide area, should be robust to all but major environmental perturbations. There is some evidence to suggest that low surface temperatures favour hake recruitment, or at least that the recruits are more abundant and at higher densities during cool periods, e.g. in 1992 in the case of M. capensis in Namibia, and in 1987 for M. paradoxus in the southern Benguela. Also, there has been a clear positive correlation between monthly catch rates and SST in Namibia in certain years (e.g. 1994 through 1996), although in other years (e.g. 1993 and 1997) the correlation has been as clearly negative (Boyer et al. 1998). The reasons for these apparent connections are not understood. The response of hake to hypoxic conditions is of particular interest in Namibia, where levels over a large part of the shelf can become intolerable even to hake, possibly causing major shifts in distribution, affecting recruitment strength (Woodhead et al.1996), and causing increased mortality of juveniles and older fish if extensive and persistent enough. For example, Hamukuaya et al. (1998) found that persistent and pronounced hypoxic conditions off central and northern Namibia in 1994 displaced M. capensis offshore, subjecting them to heavy mortality from predation by larger hake and trawling. Improving understanding of the effects of temperature and oxygen fluctuations on the distribution, abundance and behaviour of hake in the Benguela region is the focus of a number of local, regional and international research efforts. Little is known about the reaction of adult horse mackerel to environmental perturbations in the Benguela, although it is believed that in Namibia, warm-water intrusions can cause the fish to move closer inshore (Klingelhoeffer 1996). This is supported by the fact that in Namibia over the past five years, there has been a clear positive correlation between seasonal trends in CPUE and surface temperature along the 200m isobath (Boyer et al. 1998), and by historical catch data, which show that there was a large-scale southward shift in the distribution of both T. trachurus capensis and T. trecae in the northern Benguela in the late 1950s/early 1960s, coinciding with the intrusion of warm, highly saline water from the north. This was followed by a northward movement from the mid 1970s, following a period of cooling. The effect of the environment on the distribution and migration of horse mackerel in the Benguela as a whole is an important trans-boundary question, inter alia for interpreting and comparing survey results in neighbouring countries. The possible effect of environmental changes on the deep-water resources of the region is totally unknown, but it could be substantial. For example, the fact that orange roughy off Namibia concentrate within a narrow temperature range and spawn on very specific sites suggests that any significant change in the near-bottom temperature could have a major impact on the distribution and perhaps the spawning process, which could severely disrupt the fishery targeting these sites. 6.3 Crustaceans There is evidence that the declines in J. lalandii lobster production in Namibia and South Africa which occurred towards the end of the 1980s were at least partly environmentally induced. In the southern Benguela, the decline resulted from reduced growth rates, whereas off Namibia it was attributed to changes in availability related to oxygen fluctuations in bottom waters, aggravated by over-fishing. Since there is relatively little longshore migration of rock lobster, and it is improbable that fishing impacted all areas simultaneously, it seems most likely that the resource responded to some large-scale change in the environment. In the south, the decline in growth rates may have been caused by a reduced biomass of ribbed mussels, or by changes in primary production and a regime shift in the benthic foodweb. 6.4 Line-caught species Relationships between linefish species and the environment in the Benguela have not been formally studied or quantified in any way. As most linefish species are predators, the effects of environmental perturbations on their distribution and abundance is likely to be secondary, through more direct effects on the abundance and distribution of their prey. 6.5 Seals and seabirds The high mortality and breeding failure of Cape fur seals at all the Namibian colonies in 1994 and 1995 was accompanied by a drastic deterioration in the condition of both pups and adults, and was clearly the result of low food availability over most of their habitat. This is confirmed by the estimates of low sardine abundance in the period between 1994 and 1996 (Fig. 14). Likewise, the major reduction in the number of Cape gannets in southern Namibia (particularly at Ichaboe Island) in the 1960s and 1970s would appear to be related to the decline of the Namibian sardine stock during this period, the sharpest decline occurring in the mid-1970s, coincident with the collapse of the fishery. The decline of the sardine stock appears to have had a similar, but somewhat less pronounced, effect on the Cape cormorant population off Namibia. 7. SOCIO-ECONOMIC IMPORTANCE Summary data on the economic value of Namibian and South African commercial fisheries in 1996 and 1997 respectively are set out in Table 2. (The information available for Angola is less detailed, and cannot be given in this form).
Table 2. Value of major industrial fisheries in Namibia and South Africa in 1996 and 1997 respectively. Namibian data from MFMR. South African data from Stuttaford (1998). Note: N$1 = R1. * Includes longline catches ** Includes midwater trawl catches The following is a more general description of the socio-economic value of the major fisheries in each of the three countries Angola The fisheries sector is very important in Angola, being the third-most important industry after oil and diamond mining. It provides nearly half of the animal protein of the country, and is an important source of employment and food to populations of the coastal regions, where it is often the only source of livelihood for the poorer population groups. Domestic consumption of fish, which was estimated at 11.1 kg per person per annum in 1994, is the highest in the region. According to the results of a survey conducted in 1992, there were at that time around 30 000 workers directly involved in activities of the fisheries sector, of which some 18 000 were involved in artisanal fisheries. The remainder were involved in industrial fisheries and public administration. In addition, it was estimated that some 5 000 persons (mainly women) were involved in informal fish trade activities. A more recent report (Delgado and Kingombo 1998) puts the number of artisanal fishermen a few years later at over 23 000, and the number of people involved in informal fish trading at between 20 000 and 30 000. Many artisanal fishermen are not able to make a living solely from fishing, and supplement their incomes by, for example, agricultural and commercial activities. At present, roughly half of the revenue from fish and fish products in Angola comes from exports, which varied in value between US$ 27 million in 1993 and US$ 46 million in 1995. Prawns are the most important product, making up 48% of the total revenue from the fishery sector in 1995, for example. The main export markets are Europe for prawns and demersal fish, African countries for small pelagic fish including horse mackerel, and Japan for tuna and crab. Although some of the resources have clearly been overexploited, others are probably still under-utilised, evidenced by the fact that, in some of the fisheries, TAC limits have often not been reached, and that total industrial catches before Independence were typically some three times higher than they are now. This is partly due to operational constraints stemming from a breakdown in infrastructure during the civil war, and the socio-political and security situation in the county at present. With greater political and economic stability, some of these resources could well contribute more to the Angolan economy than they do at present. Namibia Fisheries is the third-largest sector of the Namibian economy, behind agriculture and mining, The industrial fishery has generated more than 10% of the GDP in recent years, producing products to the value of N$ 1 374 million in 1996. Exports were valued at N$ 1 048 million in that year, making the sector the second-largest export earner behind mining. It is the second-fastest growing industry in the Namibian economy (behind tourism) with the value of production and exports now being some six times greater than at Independence. The fisheries sector is extremely important in the social economy of Namibia, particularly in Walvis Bay, which is the major fishing port and where most of the processing plants are situated. Local employment in the sector grew rapidly after Independence, with an estimated 6 000 jobs having been created between 1991 and 1994. The integration of Walvis Bay into Namibia in 1994, and the removal of the uncertainty regarding the port’s future, stimulated an influx of investment in the fishing industry and subsidiary service industries with a further growth in employment. The number of people directly employed in the fisheries sector in 1996 was about 15 000, of which some 7 500 were fishermen. Of these 43% were foreigners, mainly in the horse mackerel and tuna fisheries, a proportion that has decreased from around 66% in 1993. It has been projected that by the year 2 000, the total number of people employed in the fisheries sector will have risen to above 20 000, exceeding the original target of 15 000 set in 1992. The demersal fishery is the most valuable fishery in Namibia. In 1996 the catch had a landed value of N$ 593 million, and a final value after product beneficiation of N$ 718 million. About 90% of the catch is either sea-frozen or wetfish hake. Monkfish make up most of the remainder, with the average landed value of the catch in recent years amounting to some N$ 70 million per year (Olsen 1997). Almost the entire demersal catch is exported. The pelagic fishery is second in importance, canned sardine being the most valuable product. In recent years the total export earnings from the pelagic fishery have been around N$ 400 million per annum, except in 1996 when no fish were canned, causing exports to drop to N$ 91 million. In more normal years, canned fish, almost all of which is exported to South Africa, make up more than 90% of the export earnings of the fishery, with fishmeal contributing almost all of the remainder. The midwater trawl fishery for horse mackerel has contributed some N$ 250 million per year in exports in recent years, mostly in the form of relatively low-value frozen fish, with minor contributions from fishmeal (around 10% ) and dried-salted fish (approx. 3 % in 1996). There is little product beneficiation, the export value of the catch being typically only about 10% above the landed value. Only about 3 % of the production is consumed domestically. The deep-water fishery has made a significant contibution to the fisheries sector in recent years, with exports to the value of N$ 171 million in 1996. Orange roughy contributes more than 90% by value, and alfonsino most of the remainder. Processing (mainly the production of high-quality fillets for the USA and Japanese markets) approximately doubles the value of the catch, and is labour-intensive, providing much-needed employment in Walvis Bay. The above four industries contribute more than 90 % by product value of all of Namibia’s industrial fish production. Of the remainder, only the tuna (3%), crab (1.5%) and rock lobster fisheries (1.5%) contribute more than 1% in most years. To these must be added the recreational linefishery. Kirchner, Sakko and Barnes (in press) have estimated that between October 1997 and September 1998, some 8 800 anglers spent 173 000 days angling, and had direct expenditures of N$ 29.7 million. Value added to gross national income within the shore-angling fishery during that period was estimated at N$ 14 million. The expenditures ultimately resulted in gross national income of some N$ 3 000 per angler, or N$ 27 million in aggregate. South Africa The living marine resources of the Benguela Current form the basis of a fishing industry which supports some 26 000 people (mostly in the Western Cape), and supplies food for the whole Southern African subregion. In 1997 the South African fishing industry caught a total of 445 000 tonnes of fish, shellfish and seaweed nationwide, of which more than 90% was taken from the Benguela. The wholesale value of the total processed output in this year was estimated at R 1 953 million, with an export value of R 873 million, on a par with Namibia. Fishing is particularly important in the social economy of the Western Cape, where some entire coastal communities depend directly or indirectly on fishing for their livelihood. However, the fishing industry yields less than 1% of South Africa’s GDP. In terms of volume, the purse-seine fishery for pelagic species is the most important sector. In 1997 (a comparatively poor year), landings of pelagic fish totalled 286 000 tonnes, of which about a third was canned. Practically all of the remainder was reduced to meal. Because of the high local demand for fishmeal, and the comparatively small output of canned fish, the pelagic sector exports relatively little (export value R 31 million in 1996). The sector is entirely industrialised, the smaller vessels (some of which are privately owned) concentrating on anchovy and juvenile sardine for meal, and the larger, factory-owned vessels on adult sardine for canning. Economically, the trawl fishery is the most important sector of the South African fishing industry. Catches of hake, which amounted to 147 000 tonnes in 1997, usually contribute about 70% of the trawl catch and about 80% of its value. Horse mackerel, snoek, monkfish and kingklip are the most valuable other trawled species, together accounting on average for about 20% by landing and value of the catch. In 1997 the landed value of processed products from a total demersal trawl catch of 200 000 tonnes was R 428 million. The value of hake exports in 1997 exceeded R 300 million; about a third of the total revenue from all South African fish and shellfish exports. The fish are largely caught by trawlers operating under quotas held by a number of large companies, although in recent years a number of smaller companies and private boat-owners have entered the trawl fishery. A longline fishery from smaller vessels has also been developing, accounting for about 3% of the hake catch in 1997. The West Coast rock lobster fishery is a major export fishery in South Africa, about 75% of the catch being exported. In the 1997 season, 1 726 tonnes of rock lobster were landed from the West Coast, with a wholesale processed (mainly frozen tails) value of R 102 million. The rock lobster fishery is labour-intensive, and is an important source of employment and income in many fishing villages on the Cape West Coast. The wholesale processed value of all commercial landings of linefish in South African waters in 1997 was estimated at R 106 million, of which about half was contributed by snoek. Contributions from tuna catches in this year made up 12% of the remainder. These figures do not represent the substantial direct and indirect contribution which recreational and subsistence fishing on linefish species makes to the South African economy. A recent nationwide survey conducted between 1994 and 1996 (Brouwer et al. 1997, McGrath et al. 1997) showed that over that period there were some 3 000 registered commercial linefish boats and about 7 900 skiboats operating off the South African coastline. About 18 100 crew were employed on the commercial boats, while nearly 14 000 recreational fishermen went to sea on skiboats. They estimated furthermore that roughly 412 000 people participated in shore-based angling, and about 7 000 each in beach-seining/gill- netting (largely a subsistence fishery) and recreational spearfishing. In all, they estimated that South Africa’s linefisheries and direct support industries provide employment to over 130 000 people, and that some 20 000 households living in poverty depend on linefish catches for about 9% of their household income. They put the total contribution of linefisheries to the gross geographic product (GGP) of South African coastal provinces (Western and Northern Cape, Eastern Cape and KwaZulu-Natal) at nearly R2 200 million, which amounts to 1.3% of the GGP of those provinces. Although a significant proportion of this was caught on the South and East Coasts, it is clear that the value of the South African linefisheries in the Benguela system is out of all proportion to the product value of the catch. 8. MANAGEMENT 8.1 Policy and legal framework In all three countries of the region it is the national policy to utilise living marine resources on a sustainable basis for the benefit of the nation, and to manage them according to scientific information and principles. Ultimate responsiblity for control measures rests with the State in all three countries. Most of the primary research on fisheries resources has been done by state-run research institutes operating within Government Departments (viz. the Ministry of Fisheries in Angola, the Ministry of Fisheries and Marine Resources in Namibia, and the Department of Environmental Affairs and Tourism in South Africa). Angola The nation’s marine and inland fisheries are managed and developed in terms of the Fisheries Act, which was developed with the assistance of the FAO and promulgated in 1992. The Act covers such aspects as fisheries management (which is implemented through various Executive Decrees governing different sectors of the fishery), planning and licensing, the control of the quality and export of fish products, and surveillance and enforcement. In recent years, with the move to a market economy in Angola, and the privatisation of large State-owned companies, the State has limited its activities to the management of the resources, surveillance, support of development and the creation of infrastructure. The broad national policy regarding fisheries development centres around the stengthening of regulatory and management capabilities of the Government, the development of small-scale fisheries, developing and increasing the participation of the national fleet in industrial fisheries, the rehabilitation of land-based industries with an emphasis on frozen, salted and canned products, and the improvement of the quality and distribution of fish for domestic and export markets. In terms of this Policy, the State is encouraging conversion of present licencing agreements for foreign fishing into joint ventures involving local vessels and Angolan entrepeneurs. Research is carried out by IIP, the Instituto de Investigaçăo Pesqueira (Institute of Fisheries Research), and IPA, the Instituto de Desenvolvimento Pesca Artesanal (Institute for Development of Artisanal Fisheries), both of which fall under the Ministry of Fisheries and have headquarters in Luanda, with smaller regional laboratories along the coast. IIP annually submits a document on the current state of the fisheries resources in the Angolan EEZ and recommendations on TACs and other control measures to the Ministry of Fisheries, and maintains a corps of some 120 observers for monitoring catches. As part of the national surveillance system, Angola is already implementing a VMS for fishing vessels, which is one of the more advanced in the region. (CHECK_FONTES) The Ministry of Fisheries receives support for research and development from donor agencies such as the Swedish International Development Agency (SIDA) and the Norwegian Agency for Development Co-operation (NORAD), and from fishing agreements with the European Union (EU). Cooperation with international bodies (such as the FAO) and various donor agencies and overseas laboratories (such as the Institute for Marine Research in Bergen and the Portuguese Institute of Marine Research in Lisbon) in the development and management of Angola’s fisheries is seen as very important. The development of links with other countries in the region (particularly Namibia) is also regarded as important, as is participation in regional marine science programmes such as BENEFIT (see Section 8.3), whose training and infrastructure-building goals are seen as being particularly pertinent to the needs of the country. Namibia In Namibia, a 200 nautical mile Exclusive Economic Zone was declared on Independence in 1990, followed by the promulgation of a new Sea Fisheries Act in 1992, and the introduction of a new national policy on exploitation rights and quota allocation in 1993. A major emphasis has been placed on Namibianization of all sectors of the fishing industry and the building up of local research and management capacity. Fisheries research is conducted within the Directorate of Resource Management of the Ministry of Fisheries and Marine Resources (MFMR), by the National Marine Information and Research Centre (NatMIRC) in Swakopmund and the Lüderitz Research Centre. Scientific recommendations for the harvesting of all resources except seals are presented to the Namibian Sea Fishery Advisory Council, which makes recommendations to the Minister of Fisheries and Marine Resources after considering socio-economic factors and the industry’s perception of the state of the resource. The Council also advises on the allocation of a research fund derived from levies on catches. The Minister, after consultation with a Fisheries Management Committee within the Ministry, submits TAC recommendations to Cabinet for final endorsement. Legislation is effectively implemented. All fish must be off-loaded under inspection at either Walvis Bay or Lüderitz, and a fisheries observer trained in basic biological sampling accompanies all vessels large enough to carry extra personnel. Surveillance is carried out by patrol vessels and aircraft, and a satellite vessel-monitoring system is being investigated. In addition to her national responsibilities, Namibia has established a SADC Sector Coordinating Unit within the MFMR to discharge her responsibility as Sector Coordinator for Marine Fisheries and Resources for the SADC. South Africa Until very recently, management of South Africa’s living marine resources was carried out in terms of the Sea Fisheries Act of 1988. TACs and other control measures were decided upon by the responsible Minister (most recently the Minister of Environmental Affairs and Tourism), acting on advice from his Department and a Sea Fisheries Advisory Committee (SFAC), which received input inter alia from the Department’s Chief Directorate of Sea Fisheries. The SFAC also made recommendations on the allocation of the Sea Fishery Fund, a fund derived from levies on fish catches that was used to support research and development activities. Quotas were awarded by an independent Quota Board. A new Act (the Marine Living Resources Act of 1998) has recently been promulgated. It includes in its objectives the achievement of broad and accountable participation in decision-making processes, and the restructuring of the fishing industry to redress historical imbalances and achieve equity within the industry. The SFAC has been replaced by a Consultative Advisory Forum (CAF), which is responsible for advising the Minister of Environmental Affairs and Tourism on management and development of the fishing industry (including the setting of TACs), research direction and allocation of a Marine Living Resources Fund, which replaces the former Sea Fishery Fund. The new Fund receives income from levies, licences, penalties and other sources, which permits its disbursement to spheres of fisheries management (e.g. administration, compliance) other than only research and development. The Minister is ultimately responsible for deciding upon TACs and other control measures, and for allocating quotas on advice from his Department. Implementation of fisheries regulations is still carried out by the Department, with assistance where necessary from the South African Navy and the Police Unit for Coastal Patrols. A VMS system to assist in monitoring the movements and activities of fishing vessels is currently being tested. 8.2 Research and management capacity Local institutions In Angola, accommodation available for marine science and technology is generally adequate, particularly at IIP headquarters in Luanda. However, the laboratories are not well equipped, and the support infrastructure (technical services, communications systems, computing and library facilities etc.) is inadequate to service the needs of the Institute. The Institute’s research vessel Goa is poorly equipped and is at present not operational, making the Institute totally reliant on foreign research vessels (particularly Dr Fridtjof Nansen) for research cruises in Angolan waters. Although the Institute employs a number of research staff with post-graduate degrees in marine science from overseas universities, most are graduates of the Agostino Neto University in Luanda, where no courses in marine science subjects are provided. Consequently there is an acute shortage of professional knowledge, both in terms of numbers and skills, which is only being partly overcome by post-graduate training within the Institute and abroad. The same is true of technical support personnel. NatMIRC in Swakopmund, Namibia, currently employs a research staff of some 42 scientists, technicians and assistants, and the Lüderitz Research Centre, which is responsible for research on local resources in southern Namibia, about 10. NatMIRC’s office, laboratory, library and meeting facilities are new and excellent, and the Lüderitz facilities are even newer. NatMIRC possesses a range of reasonably modern equipment, and there is a public aquarium within the building to increase public awareness of marine issues. The Ministry operates a 47 - m research stern trawler R. V. Welwitschia (scientific capacity 9) which is relatively new and well-equipped for resource and environmental surveys in local waters. All acoustic surveys on pelagic fish are now done on this vessel, but trawl surveys are still done on Dr Fridtjof Nansen and commercial trawlers because of limitations in Welwitschia’s bottom-trawling capability. The Ministry’s scientific staff are generally well qualified, but have limited experience. On appointment few have specific training as marine scientists, and most undergo further training through studying for post-graduate degrees at South African or overseas universities and the attendance of courses and specialist workshops locally or abroad. Training and research support is also received from donors and from foreign consultants attached to or engaged by the Ministry for varying periods. Nonetheless, there are staff limitations, a particular practical one being the shortage of qualified technicians to maintain and develop the specialised equipment needed for research. Namibian institutions which are involved in marine science education, or which have the potential to become involved, are the University of Namibia (UNAM) in Windhoek, through a new course in natural resources which includes marine science subjects, and the Polytechnic of Namibia, through its nature conservation diploma. Through the BENEFIT Training Programme, inter alia, ways are being sought to strengthen the ties between NatMIRC and these institutions and other tertiary education bodies in the region. In South Africa, statutory responsibility for advising on the state and management of marine living resources, and for carrying out the necessary research in order to do so, resides with the Department of Environmental Affairs and Tourism. Research was until very recently carried out by the Sea Fisheries Research Institute (SFRI) in Cape Town, which resorted under the Department’s Chief Directorate of Sea Fisheries. The Institute had an establishment of some 150 scientific and technical staff, who conducted research on all aspects of marine science, including resource assessment, physical, chemical and biological oceanography and equipment and gear development (including electronics). The Institute itself, as well as the Chief Directorate it served, has now been restructured to meet new challenges in resource management in the country. The scientific component is now split among three resource-orientated directorates within a new Chief Directorate of Marine and Coastal Management (M&CM), the scientific establishment becoming more involved in resource management issues in an attempt to strengthen the whole Chief Directorate where it is needed most. Scientists will still however have leading roles in the new structure, within a matrix-like system which is being developed to ensure continuation of the strong scientific ethic already in place. M&CM has a number of research vessels, the largest of which are the two research stern-trawlers, R. S. Africana (78m, with a capacity for 19 scientists) and the 52m-long R. S. Algoa, which has a capacity for 13 scientists. Both vessels are excellent platforms for multi-disciplinary research, and are relatively well equipped, although some of the equipment requires updating or replacement. In recent years the vessels have been underutilised due to staffing and funding problems, and maintenance problems are increasing. M&CM has good workshop and library facilities, and possesses a wide range of oceanographic and survey equipment (some of which was developed in-house), as well as a newly-built research aquarium of world-class standard. The Department also publishes the prestigious South African Journal of Marine Science, edited by M&CM staff, which has a high current ranking in the international Science Citation Index, and in which much of the research work in the Benguela has been published. To date 20 volumes have been published, dating back to 1983. Other South African institutions actively involved in research in the Benguela are the University of Cape Town (Departments of Oceanography, Zoology, Applied Mathematics and Statistical Sciences), the University of the Western Cape (Departments of Zoology and Botany), and to a lesser extent, the South African Museum in Cape Town (Taxonomy), the University of Port Elizabeth (Departments of Zoology and Oceanography), Rhodes University in Grahamstown (Department of Ichthyology and Fisheries Science), the Port Elizabeth Museum and the J.L.B. Smith Institute of Ichthyology in Grahamstown, a national facility of the National Research Foundation. Technical training in oceanography is offered by the Cape Technikon in Cape Town, which runs a 3-year diploma in oceanography, with practical training and lecturing by M&CM staff. These institutions have been an important source of professional and technical staff for South African marine research institutions, and strong links have been developed between them and the State, for example through the Benguela Ecology Programme (BEP); a highly successful collaborative research venture between the former SFRI and several of the universities (particularly UCT) which was started in 1981 and is still running (see assessment by Field, 1996). In South Africa, a national oceanographic data base for physical and chemical data is maintained by the CSIR’s (Council for Scientific and Industrial Research) South African Data Centre for Oceanography (SADCO) in Stellenbosch. High-resolution raw and partly-processed thermal and ocean colour imagery can be purchased from the CSIR’s Satellite Applications Centre (SAC) in Haartebeeshoek, which maintains an archive of NOAA AVHRR imagery dating back to 1984. These Centres are capable of serving the needs of much of the region, although there are inadequacies such as incomplete satellite cover of northern Angola and the lack of biological information within SADCO. Donor assistance Marine research in the Benguela has been, and continues to be, supported by donations and other assistance from foreign governments such as Norway, Germany, Iceland, Sweden, Denmark, France, the United Kingdom, Spain and Japan, plus the European Union. Assistance is also being received from international organisations such as the FAO. Through the Nansen Programme, sponsored by the Norwegian Agency for Development Co-operation (NORAD), the research vessel Dr Fridtjof Nansen has been active in the South-East Atlantic since her commissioning in 1994, and is to remain in the region until at least 2002 to assist in the national programmes of Namibia and Angola, and the regional BENEFIT Programme. Dr Fridtjof Nansen is a 57-m multi-purpose vessel excellently equipped for stock assessment surveys and studies on fishing gear performance and fish behaviour. She replaces her predecessor of the same name which carried out stock assessment and environmental surveys in Namibian waters from Independence in 1990, and off Angola since 1985, in terms of an earlier phase of the Programme. The new phase, which was launched in 1993, places greater emphasis on training and capacity-building in fisheries research and management, and has recently been expanded to include the strengthening of local fisheries institutions, particularly in Namibia and Angola. With the transition to democratic rule in South Africa in 1994, the Nansen Programme established links with marine research institutions there (particularly Sea Fisheries, now M&CM), and has endeavoured to strengthen regional co-operation in fisheries research in the Benguela. In line with this, the Nansen Programme has actively supported the BENEFIT initiative since its conception, and is now a major provider of financial and material support to BENEFIT, the latter in the form of ship’s time on Dr Fridtjof Nansen and assistance from Norwegian scientists on the staff of the Institute for Marine Research (IMR), Bergen. The German government, through GTZ, the Deutsche Gesellschaft für Technische Zusammenarbeit (German Organisation for Technical Co-operation) has supported marine environmental research and monitoring and training in Namibia since 1993, and has also been an active supporter of the BENEFIT Programme throughout its development stages. It has recently committed funds to support BENEFIT in a number of ways over the next three years, including the funding of a number of regional environmental research activities aimed at improving understanding of the impact of the environment on the major resources of the region. In addition, the German government has funded a combined research and training cruise to the region on a chartered vessel (Petyr Kottsov) as a contribution to BENEFIT, and has promoted collaboration between regional and German scientists, mainly from the Institüt für Ostseeforschung, Warnemunde (IOW) and the Centre for Tropical Marine Ecology (ZMT), Bremen; collaboration which is expected to continue under the umbrella of BENEFIT. In Angola, the Swedish and Danish International Development Agencies (SIDA and DANIDA) have in the past given considerable assistance in building infrastructure for fisheries research and development, with a particular recent emphasis on artisanal fisheries, while in Namibia, the Icelandic International Development Agency (ICEIDA) has provided assistance to the Ministry of Fisheries and Marine Resources (MFMR), mainly in the operation of Namibian research vessels and the training of officers and crew. ICEIDA has also supported the SADC Fisheries Sector Co-ordinating Unit in Windhoek. DIFD, the Department for International Development of the United Kingdom (formerly ODA, the Overseas Development Agency), has developed a fisheries information system for MFMR, and is currently investigating ways of improving the collection of fisheries statistics in the whole of the Southern African region. The Japanese government built and donated R. V. Welwitschia to the Namibian government, and made a small vessel (R. V. Matsuyama Maru) and researchers available for specific research projects for a two-year period. Namibia has also received training assistance from a number of countries and donor agencies and the FAO, the latter in the form of stock assessment courses and advice by expert consultants. The FAO has also actively supported courses in South Africa. The French government is currently supporting a bilateral study with South Africa, aimed at providing new tools and information for the regional assessment of pelagic fish resources in the Benguela. The project, code-named VIBES (Variability of exploited pelagic fish resources in the Benguela ecosystem in relation to Environment and Spatial aspects) involves collaboration between the French Research Institute for Development Co-operation (formerly ORSTOM, now IRD), M&CM, UCT and other universities and research institutes in South Africa and France. It is to be extended and expanded into the region through affiliation with the BENEFIT Programme. The region as a whole is also to receive assistance through a three-year European Union – funded international collaborative project on environmentlal conditions and fluctuations in the distribution of small pelagic fish in the Benguela (code-named ENVIFISH). The partners are Angola, Namibia, South Africa, Germany, Norway, Portugal, the United Kingdom, the European Union Joint Research Centre in Ispra, Italy, and the FAO. ENVIFISH will be closely linked to both BENEFIT and VIBES. 8.3 International and regional agreements and conventions FAO Code of Conduct for Responsible Fishing Angola and Namibia are signatories to this Agreement. South Africa is yet to sign, but has agreed in the interim to abide by its provisions. United Nations Convention on the Law of the Sea (UNCLOS) - South East Atlantic Fisheries Organisation (SEAFO) Angola, Namibia and South Africa have all ratified UNCLOS and have voted in favour of its Convention on Transboundary and Highly Migratory Stocks, and the United Nations Implementing Agreement (UIA) relating thereto. Subject to that Agreement, Angola, Namibia and South Africa, along with the United Kingdom (acting on behalf of its Dependencies; Ascension Island, St. Helena and Tristan da Cunha), have formulated the South East Atlantic Fisheries Organisation (SEAFO) for the conservation and management of straddling and High Seas stocks in the South-east Atlantic. Other parties which have expressed interest in SEAFO are the European Union, Japan, Norway, Russia, Ukraine and the USA. Negotiations on this Agreement are far advanced, and will ultimately lead to regional arrangements for the management of straddling and High Seas stocks in the region. This is likely to be the first Agreement concluded under the UIA. International Commission for the Conservation of Atlantic tunas (ICCAT) Angola and South Africa are both long-standing members of ICCAT and Namibia is about to join the organisation. The BENguela Environment Fisheries Interaction and Training (BENEFIT) Programme BENEFIT is a regional marine research and training programme involving Angola, Namibia and South Africa, with financial and other assistance from a number of Northern Hemisphere countries which have recently been active in marine research and training in the Benguela region, such as Norway and Germany, and the African Development Bank. The Programme is aimed at improving knowledge and understanding of the dynamics of key commercial stocks in the Benguela (primarily hakes, horse mackerels, small pelagic fish and crustaceans) and of linkages between environmental processes and stock dynamics, with the broad objective of improving management of these resources. BENEFIT has the full support of the Angolan, Namibian and South African governments, and of SADC, all of which are represented on a Policy Committee which guides the Programme through a network of Committees and Working Groups, on each of which all three countries are represented. International scientific guidance is provided by a Scientific Advisory Panel, on which France, Germany, Norway, South Africa, USA and the United Kingdom are represented. BENEFIT has been conceived as a ten-year programme, and will operate in terms of a locally-developed Science Plan (Shannon and Hampton 1997) which inter alia identifies the broad scientific and training problems of the participating countries, and stresses the need for a regional approach to their solution. The Programme is now underway, and the first research projects (all of which will be regional in nature) started in 1999 with assistance from the Norwegian and German governments (through NORAD and GTZ respectively). The specific training needs of the region are being identified, and contacts between local fisheries research organisations and tertiary education establishments are being strengthened with a view to developing an integrated regional training programme for BENEFIT. In addition, in mid-1999 the African Development Bank sponsored a 40-day BENEFIT training cruise in the region on R. S. Africana, which provided training in marine science to other African countries in addition to the BENEFIT partners. It is envisaged that there will be close links between the BENEFIT and BCLME Programmes which, although differing in emphasis and scope, will be mutually complementary. 8.4 Management measures for major resources Pelagic fisheries In Angola, sardinellas, horse mackerels and sardine have been assessed acoustically since 1985 from first the original, and then the new Dr Fridtjof Nansen (see estimates in Figs. 14, 17 and 20). In the absence of reliable catch statistics for these species, IIP has based management recommendations solely on trends in the survey estimates. The fisheries are managed by TAC, with no distinction between the two sardinella or two horse mackerel species. At a recent international workshop on the management of small pelagic fish in Angola, Congo and Gabon, attempts were made to estimate MSY for the region’s sardinella and T. trecae stocks using surplus production models for the former and an age-structured model for the latter. CPUE indices needed in these models were derived indirectly from the acoustic survey data and information on total catches. The sardinella models (which were considered to be more reliable than the horse mackerel model) placed the MSY at more than double the catch in recent years (around 60 000 tonnes) suggesting that these species are curently underexploited. In contrast, the horse mackerel model suggested that the current level of catch of around 60 000 tonnes per annum is approaching the species’ sustainable level. The Workshop emphasised, inter alia, the need for reliable direct CPUE indices for all the species considered, the collection of fleet and country-specific length frequency data, and the need for an effective monitoring, control and surveillance system in Angola and its northern neighbours. Prior to Independence in 1990, the management of pelagic fish in Namibia (primarily sardine and anchovy), was based on stock assessments by South African scientists, derived from CPUE indices, aerial and acoustic surveys and VPA of commercial catch data, supported by trends in the diets and breeding success of predators, and in guano production. Since Independence there have only been TAC restrictions on sardine and, in recent years, juvenile horse mackerel. Anchovy catches are however restricted somewhat by a closed season and limits on the by-catch of sardine. Recommendations on the sardine TAC have been based on acoustic/midwater trawl surveys conducted by first the old, then the new Dr Fridtjof Nansen, and the MFMR research vessels Benguela and Welwitschia. Extensive use is made of fishing vessels as scouts to find shoal groups and check that fish have not been missed close inshore or outside the surveyed area. Attempts have also been made by NatMIRC to deduce population trends in sardine from VPA and length-based cohort analysis of commercial data, but this work has been severely hampered by the unreliability of the ageing techniques used and insufficient information on population parameters. Consequently, the results have not yet been considered in management recommendations. At present, the recommended TAC for the forthcoming season is taken as 18 % of the survey estimate at the end of the previous fishing season, with subsequent adjustments if surveys during the season indicate unusually high or low recruitment, growth or mortality. This procedure has enabled management authorities to react to major resource fluctuations but, as has been emphasised at a recent international workshop on research and management of the Namibian sardine (Anon. 1997b), there is a pressing need for more rigorous stock assessment modelling, using all appropriate data, as the foundation for management decisions. In South Africa, the most important pelagic resources (i.e sardine, anchovy and round herring) have been routinely surveyed acoustically since the mid 1980s, with surveys of recruitment in winter and spawning biomass in summer (e.g. Hampton 1992, 1996). Between 1984 and 1993, the anchovy spawning biomass was also estimated annually by egg surveys, using the daily egg-production method; a method which is now being considered for sardine. The survey estimates of sardine and anchovy spawning biomass and recruitment strength, and of the precision of these estimates, are used together with (in the case of sardine) estimates of the population age structure from commercial data, to model the risk to the stocks of various harvesting strategies, and hence to recommend TACs for sardine and anchovy. This process is carried out through a Pelagic Working Group, with input from Sea Fisheries survey personnel, modellers from the Department of Applied Mathematics, UCT and M&CM, environmentalists and, on occasion, the pelagic fishing industry. The current strategy is to manage the sardine and anchovy fisheries together and interactively in such a way as to optimise the sardine catches, while heeding the needs of the anchovy fishermen, who frequently catch juvenile sardine as a by-catch. There are at present no direct restrictions on the round herring fishery, because the resource is not thought to be threatened by the comparatively low levels of catch (typically less than 50 000 tonnes per year from a stock which has consistently been estimated acoustically at about 1 million tonnes). Trawl fisheries Hake off Angola (Merluccius polli and, in the extreme south, M. capensis), have been investigated in the course of various bottom-trawl surveys conducted by R. V. Goa between 1970 and 1992, the old and new Dr Fridtjof Nansen between 1984 and the present, and recently, by chartered fishing vessels. As a part of these studies, the other major groups of demersal species in Angolan waters (dentex, croakers and groupers) have also been investigated. In the absence of reliable fisheries statistics for any of these species in Angolan waters, stock assessments and TAC recommendations have been based on trends in the survey estimates using holistic models. With improvement of commercial catch information, it will be possible to incorporate CPUE data into the analysis. Separate TACs are set for the two hake species and for different groups of other demersal fish. Other forms of control include effort limitation, the prohibition of trawling close to the coast, and minimum size limits. Between 1975 and 1989, the assessment and management of Namibian hake stocks was carried out under the auspices of ICSEAF. Various surplus production models based on catch and effort data from the Soviet and Spanish fleets were used in the assessments. The fishery was managed by mesh regulations and limits on the TAC, which was apportioned between nations according to their historic interest and performance in the fishery. Since Namibia’s declaration of an EEZ in 1990, and the subsequent withdrawal of foreign fleets, the hake TAC has been based on biomass estimates obtained from Dr Fridtjof Nansen bottom trawl surveys, in which Norwegian and Namibian staff participate. To these estimates are added acoustic estimates of hake off the bottom. The surveys produce estimates of the fishable (> 35 cm) and non-fishable (<35 cm) components of the population for both M. capensis and M. paradoxus. Trends in the survey estimates of the adult stock and of recruitment strength are combined with CPUE indices of trends in the adult stock to recommend annual adjustments in the TAC. Previously, the recommendation was set at 20% of the estimated fishable stock, but a new Operational Management Procedure, in which the TAC is adjusted according to the average change in the survey and CPUE indices for the previous 5 years (Butterworth and Geromont 1997), has recently been recommended as an interim measure until the question of whether the survey estimates can be treated as absolute has been resolved. (Treating them as absolute indicates that the resource is currently heavily depleted, in conflict with a number of different production models, which indicate the opposite – Anon. 1997c. The discrepancy is resolved if the survey estimates are treated as relative rather than absolute). M. capensis and M. paradoxus in South African waters are assessed as one for management purposes, using commercial data in a locally-developed dynamic Schaefer-form production model, which since 1989 has included multiple CPUE series, and is tuned by data from research swept-area surveys using bottom trawls. Recently, the Fox form of production model has been applied, and advances made in incorporating age structure for the hake stock on the South Coast, where the trawl and linefisheries select different age components of the population (Geromont et al. 1999). The assessments are carried out by M&CM, with modelling assistance from the Department of Applied Mathematics, UCT. The group together makes annual TAC recommendations through a Demersal Working Group, which liaises with the demersal industry when and where appropriate. The survey estimates used in the assessments are obtained from annual swept-area bottom trawl surveys of the West and South Coasts from R. S. Africana, using pseudo-random depth-stratified sampling. The estimates are treated as relative because of difficulty in quantifying catchability coefficients. Research is currently directed at refining the assessments by using Generalised Linear Models to improve the validity of the CPUE time series, investigating the effect of wind stress on commercial catch rates, and disaggregating the commercial catch by species so that a species-specific VPA can be performed. Since 1984, assessment of Cape and Cunene horse mackerel in Angolan waters has mainly been done from acoustic surveys conducted by the old and the new Dr Fridtjof Nansen, most of them as extensions to Namibian surveys. Information relevant to stock assessment has also been collected during R. V. Goa cruises between 1972 and 1992, as well as from collaborative resource studies with the Atlantic Research Institute for Fisheries and Oceanography (ATLANTNIRO) of the former USSR. As with other commercial species in Angola, catch statistics on the horse mackerel fishery are inadequate for assessment and management of the fishery, and TACs (for the two species combined) are based on trends in the survey estimates . In Namibia, adult horse mackerel were assessed and managed from 1980 to 1989 according to TACs set by ICSEAF agreements, based on Schaefer and Fox surplus production models applied to catch data from the international midwater trawl fishery. No distinction was made between Cape and Cunene horse mackerel, although the former dominated in the catches. TACs were allocated between interested nations by ICSEAF in a similar manner to the hake TACs. Since 1990, when the fishery came under Namibian control, TACs for the midwater trawl fishery have been based on MFMR recommendations, made according to trends in acoustic survey estimates, recently supported by length-based and age-based VPA estimates, obtained using commercial catch data. The reliability of horse mackerel ageing techniques needs to be substantially improved before the age-based VPA estimates can be used with greater confidence. In South Africa, the lack of a reliable age-structured catch and CPUE data series has hampered attempts at producing reliable stock assessments of T. trachurus capensis. A surplus production model, based on CPUE indices, egg-density data and abundance indices from direct surveys, was used to assess the resource on the South Coast between 1989 and 1991 and recommend TACs through the Demersal Working Group. The first TAC (30 000 tonnes) was set in 1990. These assessments were discontinued in 1991 when withdrawal of the Japanese vessels terminated the CPUE time series. Since 1993 a Beddington and Cooke-type yield-per-recruit model, based primarily on bottom-trawl survey estimates has been used to set a precautionary catch limit. The estimates are obtained from surveys on Africana which are biased to a highly variable degree because of spatial and temporal variations in the availability of horse mackerel to bottom trawls. To reduce this bias, acoustic techniques are now being introduced to estimate the portion of the population on the South Coast which is inaccesible to the bottom trawl (Barange et al. 1998). Management of the deep-water fisheries off Namibia is based on TAC recommendations from the Namibian Deep Water Fisheries Working Group to the Namibian Sea Fishery Advisory Council. The Working Group consists of MFMR scientists and industry representatives, and receives input from a number of foreign scientific and industry consultants. For orange roughy, management is based on a population model which uses acoustic and swept-area survey estimates of abundance on the three main grounds, and swept-area estimates of abundance on these grounds from commercial catches throughout the season, to recommend TACs for each of the grounds. The current policy for the fishery is to have a fishing-down period at a constant catch, followed by a gradual reduction in catches to a level likely to provide MSY. No TACs have been recommended for the Alfonsino fishery because of the low level of catch. This situation will be reviewed should the annual catch rise above 2 000 tonnes. The monkfish fishery in Namibia is at present managed by effort control, mesh-size limits and by-catch penalties on catches of monkfish by hake trawlers, which in recent years have made up more than 30% of the monkfish catch. Recently there has been pressure to move to a catch-limited control system. Crustacean fisheries West Coast rock lobster resources in Namibia and South Africa are assessed and managed according to various population models. In Namibia, a Schaefer surplus production model based on annual catch and effort data is used to recommend TACs and minimum size limits, with the assumption that the stock at the start of the time series (1958) was 40% of pristine. Fishing is controlled by limits on the TAC per area, a prohibited area and closed season, minimum size and bag limits, and various restrictions on catch methods. Stock assessment-related research being conducted by MFMR staff in Lüderitz includes investigation into the effects of migrations on sex ratios, the estimation of growth rates through tagging studies, and studies on the effect of environmental conditions (particularly oxygen levels) on CPUE indices. In South Africa, the J. lalandii resource has been assessed and managed since 1992/93 through a size-based population model which uses data on growth rates, size structure and sex ratios in catches, CPUE and total landings. Results from a Fisheries-Independent Monitoring Survey have also been used. The model, which has been modified and updated a number of times since its inception, is used to make recommendations on TACs for the commercial fishery, and on a minimum size limit. The catch is also controlled by a closed season, prohibited areas, a bag limit and other restrictions on recreational fishermen. Very recently, a new Operational Management Procedure has been introduced to facilitate TAC recommendations. The input data required are the TAC from the previous year, and three of the indices of resource status (averaged over the three previous seasons), which were used previously in the size-based model (Anon. 1998b). The management of deep-sea red crab in Namibia is based on length-based cohort analysis and prediction models, adapted to fit the growth dynamics of the species, using growth rates established by tagging (Le Roux 1997). The models are used to project future stock size as a function of catch, from which TACs are recommended. The catch is also controlled by limits on minimum size and the prohibition of fishing inside the 400m isobath. Management of the resource in Angola is based on CPUE trends and estimates of MSY, using estimates of natural mortality from a number of different sources. Catches are controlled by TAC, effort control, prohibition of fishing inside 500m to protect juveniles and immature females, a minimum size limit and limitation of the crab by-catch in the prawn fishery. The deep-water prawn fishery off Angola is managed on the basis of trawl survey estimates and CPUE trends, which are used in a simplified Beverton and Holt model to recommend TACs for rose prawn and striped red prawn. Other forms of control include the prohibition of fishing within certain inshore areas to protect juveniles, a closed season and effort limitation /reduction. Linefisheries There are effectively no restrictions on catches in the large artisanal fishery in Angola, partly because of the difficulty in enforcing regulations. Consideration has however been given to protecting the interests of small-scale fishermen by prohibiting trawling close to the coast, which can severely disrupt small-scale fishing operations. The issue has not been resolved and remains a source of conflict between industrial, semi-industrial and artisanal fishermen in Angola. Management of the tuna fisheries in Angola and South Africa is carried out in line with ICCAT regulations. Although not yet a member of ICCAT, Namibia is also following these regulations, and implements effort control over both national and foreign vessels. The commercial line fishery for snoek and angling species in Namibia is at present unrestricted, but recreational catches of angling species are controlled by closed areas and bag limits. In South Africa there is a comprehensive suite of linefish management regulations developed by the National Marine Linefish Committee, which was set up in 1984. These include regulations on the licensing and number of permits for commercial fishing boats, bag limits by species category for all recreational and part-time commercial fishermen, closed seasons for certain species, and minimum size limits for the most important ones. After 1985 the Committee was superseded by an independent body; the South African Marine Linefish Management Association (SAMLMA), which has provided advice on modifications of the original measures. The new Marine Living Resources Act of 1998 retains most of the past measures, with some changes in the commercial permit system and the institution of a "Subsistence" fishing category, in which fishermen are subject to bag limits but permitted to sell their catches. Recreational fishermen are also now obliged for the first time to obtain an angling permit, although as before, will not be permitted to sell their catches. Continuing reduction in the abundance of most linefish species has led to the development of a holistic mangement protocol for linefish, which is currently being taken forward. Seals The Namibian seal harvest is primarily controlled through limitations on the annual TAC, with separate allowances for pups and bulls and for the different colonies. TAC recommendations are based on aerial censuses and estimates of biological parameters for the population (fecundity rate, mortality of pups and adults, sex ratios etc.), which are used in a deterministic, age-structured model of the female component of the population to predict ideal harvesting levels for maintaining sustainable yields. The total seal harvest in recent years (which has not always reached the TAC) has varied between 17 000 animals in 1991 and 38 000 animals in 1994, with pups contributing about 80% of the harvest in all years. 9. RESEARCH GAPS AND THREATS TO MANAGEMENT There are a number of broad gaps in scientific understanding of the dynamics of the Benguela Current’s marine resources which inhibit rational, optimal management of these resources in all three countries. The major problems have been identified in the Draft BENEFIT Science and Implementation Plan (Shannon and Hampton 1996), and are the focus of a recently-developed framework for resources research within the Programme (Anon. 1998d). Briefly, they can be summarised as:
Those of the above which are trans-boundary problems in resource management are summarised in Table 3. Included in the Table is an indication of the immediate and root cause of each of the problems listed. It will be noted that the root cause of many of the problems is the lack of regional agreements and structures for research and management of shared resources, and the shortage of manpower and funds to undertake trans-boundary surveys and other trans-boundary research activities. Table 3. Immediate and root causes of major trans-boundary problems in the manage-ment of the region’s marine living resources
In addition to these general scientific problems (which are not unique to the Benguela), there are particular scientific and operational problems and threats to management in each country, which differ according to the nature of the fisheries and the economic realities and research and management capacity in each country. In Angola, the resources and their environment have been significantly less studied than elsewhere in the Benguela, and the history of fisheries research is too short to have provided a long time-series of observations and a strong scientific foundation for the proper analysis of trends in population size (Neto 1998). There are limited national data for long-term retrospective analyses of major fluctuations in the marine ecosystem, large deficiencies in the understanding of fundamental life history characteristics (e.g. stock delineation, location of spawning grounds, distribution of ichthyoplankton, nursery grounds, migration patterns) of commercially important stocks, and no population models which can be used to evaluate management options. Partly because of the large number of remote landing points, and a small corps of compliance officials, catch and effort statistics for many of Angola’s fisheries tend to be unreliable, making it difficult to implement even basic management measures. Research capacity is limited because of the small number of people involved, the lack of appropriate tertiary education in fisheries science in the country, and the severe macroeconomic problems in the country resulting from the protracted civil war and resultant breakdown of services. There is clearly a desperate need in Angola for education and training in fisheries science and resource management at all levels, but this can only proceed if there is stability, and basic services and infrastucture are in place. In Namibia, there are fairly reliable catch statistics for all of the exploited species, and control measures are effectively implemented. The greatest general scientific problem is the development of rigorous methods of assessing sardine, hake and horse mackerel biomass from survey and other information, and the building of these assessments into formal, testable management procedures which take assessment errors into account. In view of the dramatic effects which the major environmental perturbations of the mid 1990s had on the abundance and distribution of Namibian resources (particularly sardine and their predators), a strong need has been perceived to improve understanding of the effects of the environment on the country’s marine resources, particularly with a view to predicting recruitment and anticipating changes in distribution. The major operational constraint is a severe shortage of scientific and technical staff within the Ministry of Fisheries and Marine Resources for the large number of resources that have to be studied, and the amount of effort necessary merely to maintain the current level of resource monitoring. The often prolonged absence of staff attending training courses and studying outside Namibia for higher degrees places an additional load on remaining research staff, further reducing the amount of time available for detailed analysis of results, innovative research and the publishing of results in the primary literature. Although the opportunities for local tertiary education in marine science may improve in due course, lessening the need for distance-education, this problem is likely to persist for some time. In South Africa, there are long and generally reliable time-series of catch statistics for the major exploited species, and effective exploitation control measures for most of them. The major exception is the linefishery, where attempts to limit effort have so far been ineffective. The problem is being exacerbated by technological improvements and increased capitalisation in the sector, and increasing demands on already over-exploited coastal linefish resources by a rapidly expanding subsistence sector. Although the scientific basis for fisheries management in South Africa is very much stronger than elsewhere in the region, there have been a number of developments which in recent years have weakened national capacity in marine science, and which threaten to weaken it further. State funding for marine research, both within statutory organisations and at universities is shrinking, with a consequent shortage of funds for equipment, running expenses travel and education. There is a shortage of funds to maintain, man and operate ageing research vessels, and at present little provision for replacing them. Because of these factors, increased difficulty is being experienced in maintaining even routine resource-monitoring cruises essential for recommending TACs, and there is almost no ships’ time available for developmental work and supporting (e.g. environmental) research. This jeopardises the substantial progress which has been made in the past two decades in understanding the effect of the environment on fish resources in the southern Benguela, at a time when the use of such understanding in fisheries management is being pioneered in the region. Also, in recent years there has been a weakening of scientific and research management capacity within the Department of Environment Affairs and Tourism due to loss of senior staff brought about largely by moves to reduce the size of the Public Service. The loss of expertise, and in many cases, posts, has added to the routine commitments of remaining staff, further limiting their opportunities for undertaking innovative work and otherwise pursuing their scientific careers. The weakening of state-funded capacity in marine reseach, unless compensated for by an increase in the marine research done by non-statutory bodies and through regional and international co-operative programmes, could well lead to a decline in the quantity and quality of marine science in South Africa. With the broadening of participation in the fisheries sector under the new Act, and the increased number of landing sites, monitoring is becoming increasingly difficult, which is likely to result in a decline in the reliability of data. The pressure on already hard-pressed staff within the M&CM Chief Directorate brought about by growing demands on the MCS Unit and new responsibilities to advise on quota allocation (previously the function of the Quota Board) is likely to add to staffing problems in the years ahead. 10. ACKNOWLEDGEMENTS We acknowledge generous assistance given by staff at IIP Luanda, NatMIRC, Swakopmund and M&CM, Cape Town in providing data needed for this Report. We also thank the Reprographics Section, M&CM, Cape Town for the use of figures and other artwork from their files. Drs A.I.L.Payne and P. de Barros are thanked for extensive and helpful comments on an earlier draft of the manuscript, which led to a number of improvements, and generally tightening of the text. 11. REFERENCES AND OTHER LITERATURE During the past decade several thousand scientific publications on the Benguela ecosystem and its resources have been published in the primary scientific literature, the majority of them dealing with the southern Benguela. A listing of these is beyond the scope of this document. Instead, readers are referred to Crawford et al. (1987) and references therein for a comprehensive summary of the published work on the major fish and inverterbrate resources of the Benguela up to that date, and to the book Oceans of Life off Southern Africa (Eds. A.I.L. Payne and R.J.M. Crawford, 1989, revised in 1995), which gives a popular but authoritative account of the same body of work. Much of the more recent work has been published in the South African Journal of Marine Science, particularly in the three "symposium volumes" viz. Vol. 5: The Benguela and Comparable Ecosystems (Payne et al. 1987), Vol.12: Benguela Trophic Functioning (Payne et al. 1992) and Vol. 19: Benguela Dynamics: Impacts of Variability on Shelf-Sea Environments and their Living Resources (Pillar et al. 1998). A wealth of information on the Namibian and South African fishing industries, fisheries, catches, etc. may be found in the Fishing Industry Handbook series, edited by M. Stuttaford and published annually by Marine Information CC. The references listed below give details on these publications, a short selection of other useful works, and the articles cited in the text, which have largely been restricted to work which has appeared since publication of the BENEFIT Science Plan (Shannon and Hampton 1996, 1997).
12. ACRONYMS
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||