|
SYNTHESIS AND ASSESSMENT OF INFORMATION ON THE
BENGUELA CURRENT LARGE MARINE ECOSYSTEM (BCLME)
THEMATIC REPORT NO.2
INTEGRATED OVERVIEW OF THE OCEANOGRAPHY AND ENVIRONMENTAL VARIABILITY OF THE BENGUELA CURRENT REGION
By
L.V. Shannon and M.J. O'Toole
Windhoek, Namibia November 1999

1. INTRODUCTION
The Benguela is one of four major current systems which exist at the eastern boundaries of the world oceans, and the oceanography of the region is in many respects similar to that of the Humboldt Current off Peru and Chile, the California Current off the west coast of the U.S.A. and the Canary Current off north-west Africa. These eastern boundary currents are characterized by upwelling along the coast of cold nutrient-rich water, and are important centres of plankton production which support a global reservoir of biodiversity and biomass of fish such as sardine (pilchard), anchovy and horse-mackerel and also sea birds and marine mammals.
The coastal upwelling area of the Benguela Current ecosystem extends from southern Angola along the west coast of Namibia and South Africa around the southernmost part of the continent. While the area shares many of the generic characteristics of other eastern boundary currents, it is unique in that it is bordered at both northern and southern ends by warm water systems viz. the Angola Current and Agulhas Current respectively. These equatorward and poleward boundaries are not fixed in space and in time, but are highly dynamic, and their pulsing impacts on the ecosystem as a whole and on its harvested resources. With a western boundary approximating to the 0° meridian, the Benguela thus encompasses the coastal upwelling regime, the eastern part of the South Atlantic gyre and a complex system of fronts and transition zones. In terms of the Benguela Current Large Marine Ecosystem
(BCLME) Programme, the Benguela is viewed in a broader context than is customarily defined, and includes the full extent of Angola's Exclusive Economic Zone
(EEZ), with a northern boundary at 5°S at the Angola Front. (The latter is the boundary between the BCLME and the equatorial current system.)
The earliest physical observations in the South Atlantic and Indian Oceans were those necessary for the safe and efficient passage of sailing ships along the trade routes between Europe and the East. The early Portuguese, Dutch and other navigators accordingly compiled comprehensive records of winds and currents - records which display a remarkable amount of information about the underlying physical oceanography of the region. The first published work of scientific merit was, as may be expected, of currents around the Cape of Good Hope and was compiled by James Rennel in the 18th century
(Rennel 1778). It was, however, the cruise of the
H.M.S. Challenger in the 1870s which initiated the global science of oceanography, and pioneering studies were conducted on that expedition in the Benguela region during 1873. The next half-century witnessed the age of the great oceanographic expeditions
inter alia those of the Valdivia, the Gauss, the Planet and the
Möwe. It was, however, work undertaken during the expedition of the German
Meteor in the South Atlantic between 1925 and 1927 that resulted in a quantum jump in human understanding of the oceanography of the Benguela and adjacent regions.
The scene for development of regional oceanography was set by the arrival in Cape Town of Professor J. D. F. Gilchrist in 1896. Gilchrist is regarded as the father of southern African oceanography, and he undertook a host of marine studies in Angola, Namibia, South Africa and
Moçambique. During the second half of the twentieth century it was oceanographers such as Drs T. J. Hart, R.
I.Currie, A. J. Clowes, A. H. B. De Decker, N. D. Bang, J. R. E. Lutjeharms and L. Hutchings who contributed so much to the understanding of the complex physics, chemistry and biology of the
Benguela.
This overview is a brief summary the oceanography and environmental variability of the Benguela Current Large Marine Ecosystem. As such it draws principally on the published scientific literature which now comprises several thousand authoritative articles. In preparing this overview, we have synthesised available information and ideas and have attempted, where possible and where appropriate, to simplify and to explain concepts in such a way that they will be intelligible to non-oceanographers, yet still useful to marine scientists with an interest in the Benguela system. It is not, however, a Benguela science review per se: There are a number of authoritative reviews in the international scientific literature, and these are referred to in the text, together with other key publications.
The overview commences with a discussion of the main physical features and processes in the Benguela - the bathymetry,
windfield, temperature, upwelling, currents, fronts and boundaries. Key aspects of the chemistry and chemical processes follow, including major and minor elements and all-important dissolved oxygen. The next section deals with plankton, primary and secondary production and the foodweb and carbon budget. We have also devoted several paragraphs to a discussion of environmental variability and the ecosystem consequences thereof. Finally we have provided a perspective on the various issues, problems and threats facing the
Benguela, and have identified what we believe are the major gaps in knowledge and understanding. It is hoped that the various sections collectively will provide a useful introduction to, or at least background information for, those overviews dealing with more specific aspects of Benguela resources.
2. PHYSICAL FEATURES AND PROCESSES
2.1 Bathymetry
The continental shelf along the west coast of southern Africa is variable in width and depth. It is narrow off southern Angola (20km), south of Lüderitz (75km) and off the Cape Peninsula (40km) and widest off the Orange River (180km) and in the extreme south where the Agulhas Bank extends over 200km polewards from Cape
Agulhas, the southernmost tip of Africa. The edge of the continental shelf, or shelf break as it is generally termed, lies at depths between about 200m and 500m. The position of the 200m depth contour
(isobath) is shown in Fig. 1. By global standards the Benguela continental shelf is relatively deep, and the slope and configuration of the shelf is by no means uniform. Indeed, double shelf breaks are common off the west coast of Southern Africa: for example near Walvis Bay (23°S latitude) there are inner and outer breaks beginning at depths of about 140m and 400m respectively. This is illustrated graphically in Fig. 6, which also shows a similar feature at 32°S off South Africa. Between about 31°S and 35°S several convoluted submarine canyons intersect the shelf, the best known of which is the Cape Canyon which lies approximately 80 north-west of Cape Town. The variable topography of the Benguela shelf is of particular significance for near shore circulation and for fisheries.
The continental shelf is covered by layers of sediments primarily of biological origin
(biogenic) and large areas of shelf sediments contain more than 75% calcium carbonate. Significant features of the Benguela shelf are two extensive mud belts, each about 500km long. The southern belt extends from the Orange River southwards, up to 40km wide with an average thickness of 15m, and is mainly of river origin. The northern belt which lies over the middle shelf off Namibia comprises organically rich diatomaceous oozes (originating from planktonic plants). In places the organic carbon content of these diatomaceous muds exceeds 15%!
West of the shelf break is a steep continental slope area which descends to a depth of about 5000m where it meets the abyssal plain of the South-east Atlantic Ocean. This plain comprises two large ocean basins, the Cape Basin and the Angola Basin, separated by a submarine mountain chain, the Walvis Ridge which runs from its abutment with the continental shelf at latitude 20°S (northern Namibia) in a south-westerly direction for more than 2500km towards the Mid-Atlantic Ridge. The steep continental slope and a cross-section of the Walvis Ridge is illustrated in Fig.6. As may be expected, the latter feature forms a barrier to deep circulation in the South-east Atlantic. Other prominent bathymetric features are the Agulhas Ridge which forms the southern boundary of the Cape Basin, and the Agulhas Plateau - both shown in Fig. 1, and numerous seamounts of volcanic origin, of which Vema is perhaps the best known.
2.2 Winds
Winds significantly influence the oceanography of the Benguela region on various time and space scales, ranging from basin-wide seasonal and longer period processes to local inshore events of only a few hours duration. The prevailing winds along the west coast of southern Africa are controlled by anticlockwise
(anticyclonic) motion around the South Atlantic High pressure system, the seasonal low pressure field over the land and eastward-moving cyclones which cross the southern part of the subcontinent. The South Atlantic High (Anticyclone) is part of a discontinuous belt of high pressure which encircles the southern hemisphere and is maintained throughout the year. Small seasonal differences occur. On average the anticyclone is centred at about 28°S: 8°E, and undergoes seasonal shifts, being at a more northerly and easterly position in winter than in summer. The pressure over the subcontinent alternates between a well-developed low during summer and a weak high in summer, and consequently the atmospheric pressure gradient - and hence wind - is seasonably variable. The coastal plain, much of which is arid, acts as a thermal barrier to cross-flow, and hence winds tend to be predominantly southerly
(longshore) over most of the Benguela region, being "topographically steered" along the coast. These longshore winds produce coastal upwelling which gives much of the Benguela its cool surface water characteristics (discussed in the next subsection).
The essential seasonal differences in the intensity of the upwelling-producing longshore winds is best illustrated in Fig.2. From this diagram it is evident that the principal perennial area of strong southerly winds lies near Lüderitz (27°S) with a secondary area near Cape Frio (18°S). In winter the northward shift in the atmospheric pressure systems has a strongest influence south of 31°S, where there is a relaxation of the southerly winds and a greater frequency of
westerlies. Off central Namibia wind speeds are generally lower on average, and display less seasonality. Off northern Namibia, the longshore wind is strongest during autumn and spring. North of about 15°S, the latitude of Namibe in southern Angola, the winds are much weaker than off Namibia and South Africa, although they remain longshore on average and reach maximum intensity during winter.
A common feature of the wind field during autumn and winter are "Berg" winds. These catabatic winds occur when there is a pronounced high pressure over the subcontinent, and they blow down from the central plateau across the escarpment and over the coastal plain and then seawards. They are hot, dry winds, often laden with fine particles of dust which is visible in some satellite pictures. They exert little direct physical effect on the sea, however, as being warm, they tend to blow above the cool marine atmosphere layer.
Apart from seasonal changes in the windfield, coastal winds are modulated in the southern Benguela during summer by the passage of the easterly-moving cyclones (low pressure cells) which move past the tip of the subcontinent. These result in periodic changes in winds from northerly - initially with an easterly component, before blowing from the northwest - to southerly (southwesterly to southeasterly). The latter can be quite intense and are often characterised by a "tablecloth" of cloud over Table Mountain at Cape Town. These wind relaxation-reversal-strengthening events typically occur on periods of 3 to 10 days.
Diurnal changes in coastal wind intensity and direction are common throughout most of the Benguela region north of St Helena Bay (near 33°S). These are associated with the differential heating and cooling of the sea and the adjacent land mass, typical of the classical land-sea breeze effect. Off much of Namibia coastal fog is often associated with the night time and early morning slacker winds, and tends to dissipate around noon when the southerly wind intensifies.
Readers wishing to know more about the climatology of southern Africa are referred to a definitive book on the subject by Tyson (1986), while comprehensive accounts of the winds over the ocean are contained in Nelson and Hutchings (1983), Shannon (1985), and Shannon and Nelson (1996). For comparisons of the winds and oceanography between the Benguela and the other three eastern boundary current systems, Parrish
et al (1983) and Bakun and Nelson (1991) are recommended texts.
2.3 Upwelling and surface temperature
Coastal upwelling is the process whereby cold water is brought to the surface near the coast under the influence of longshore equatorwards winds. The essential process is illustrated in Fig. 3. In simple terms, the longshore wind can be viewed as displacing warm surface water northwards and, as a consequence of the earth's rotation, offshore. This results in a drop in sea level against the coast, which serves as a non-permeable boundary. To balance the displaced water, the deeper water wells up inshore, and compensatory circulations and longshore currents over and adjacent to the continental shelf are set up. In a simple one cell system, the thermocline (layer where there is a strong vertical temperature gradient) is displaced vertically upwards, and may result in a front between the warm oceanic water and the cool upwelled water, with water moving at depth over the shelf and upwards, and sinking at the front. This is in reality an over-simplification - two or three fronts may develop with rather complex circulations in between, while the actual extent of upwelling and the intensity and direction of shelf currents will be influenced by "coastal trapped waves" - a type of internal wave within the ocean. The existence of these coastal trapped waves can result in enhanced or reduced upwelling and larger sea level changes than might simply be inferred from the wind. Nevertheless, as a general rule, the areas along the west coast of southern Africa where the southerly winds are consistently strongest are also the areas where upwelling is most pronounced. It follows, therefore, that coastal upwelling in the Benguela is neither uniform in time or in space.
The wind field, topographic features (bathemetry and land features) and orientation of the coast result in the formation of a number of areas where upwelling is more intense. The principal upwelling centre or cell is in the central Benguela in the vicinity of Lüderitz (27°S). Strong upwelling occurs there throughout the year (Stander, 1964), with some slackening during autumn, and the extensive zone characterised by cold surface water, weak stratification and high turbulence which results there appears to be an important determinant of the biology of the system - effectively dividing the Benguela into two quasi-independent subsystems (Fig. 7). There are several secondary upwelling cells viz
Cunene, northern Namibian and central Namibian cells (at approximately 18°, 20° and 24°S) and the
Namaqua, Columbine and Cape Peninsula cells (at about 31°, 33° and 34°S). The last two are seasonal, with maximum upwelling occurring between September and March, whereas off northern and central Namibia upwelling is more perennial, but with a late winter maximum. Several smaller ephemeral upwelling cells develop to the west of headlands along the south coast. Although upwelling does occur along the coast of Angola at times, it is not pronounced, and the water column remains stratified throughout the year.
In the northern Benguela peak upwelling and insolation (solar heating) are out of phase, and sea surface temperatures over the shelf follow a distinct seasonal cycle. In the south off the Cape Peninsula, maximum insolation and the upwelling season coincide, and average sea temperatures inshore vary seasonally by little more than 1°C. Viewing the Benguela in terms of a heat budget, the central zone is a major heat sink, with negative climatological sea surface temperature anomalies of 5° - 6°C in the Lüderitz vicinity. South of Africa in the area influenced by the Agulhas Current positive climatological sea surface temperature anomalies of 2°-4°C exist and the area is a major heat source for the atmosphere.
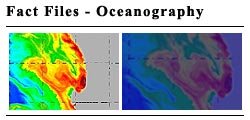
Off Angola, north of about 14°S, there is a positive offshore climatological sea surface temperature anomaly during summer. The dramatic temperature differences between surface waters of the Angolan and Agulhas Currents and those associated with upwelling off Namibia and South Africa are illustrated in Figs 4 and 8.
2.4 Water masses and general circulation
Like temperature, salinity is an important physical property of sea water, and also affects density, and density and pressure (which is approximately proportional to depth) are key parameters in terms of ocean dynamics - just as they are in the atmosphere, the only difference being that sea water is less compressible than air. Salinity is measured in "practical salinity units"
(psu) and one psu corresponds to one part per thousand or one tenth of one percent. The salinity of sea water is typically about 3.5% or 35
psu, but like temperature can vary. Salinity is influenced inter alia by fresh water input from rivers, by evaporation, precipitation, freezing of sea water and melting of sea ice.
Water masses are defined by specific temperature-salinity properties. There are a number of different water masses present off the west and south coasts of southern Africa, and their distribution and essential characteristics have been described by various authors and reviewed by Shannon (1985), Chapman and Shannon (1985) and Shannon and Nelson (1996).
The principal water masses in the Southeast Atlantic are Tropical and Subtropical Surface Waters, Thermocline Waters (comprising South Atlantic and Indian Ocean Central Water), Antarctic Intermediate Water
(AAIW), North Atlantic Deep Water (NADW) and Antarctic Bottom Water (AABW). The "core" characteristics of these are annotated in Fig. 5 which shows the typical temperature-salinity curves (relationships) for the water masses present in the South-east Atlantic. The linear part of the curve (approximately 6°C, 34.5 psu - 16°C, 35.5
psu) spans the Thermocline Water layer, and this is the water which upwells along the coast, and which constitutes, often in highly modified form, the waters present over the continental shelf in the Benguela system. From this figure it can be seen that there are quite marked differences between the Thermocline Water present in the northern and southern parts of the Benguela and true South Atlantic Thermocline Water. At the core of the layer, however, i.e. at 10° - 12°C, it is impossible to distinguish between the Thermocline Waters of different origins on the basis of temperature and salinity only. The flow of the Thermocline Water tends to be similar to that of the overlying surface water, which is discussed a little later. In a recent paper Poole and Tomczak (1999) using optimum multiparameter analysis show a clear separation between Thermocline Water in the Benguela Current system south of about 25 °S and that further north, the latter being >80% Western South Atlantic Central Water.
Thermocline Water overlies Antarctic Intermediate Water, which is formed in the Southern Ocean and which is characterised by the salinity minimum in the temperature-salinity curve. The core of the AAIW in the Benguela has a salinity in the range of 34.2 - 34.5 psu and a temperature of 4° - 5°C, and is present in the region at an average depth of 700 - 800m. In terms of volume, AAIW accounts for about 50% of the water present in the upper 1500m. The AAIW in the southern Benguela is generally much fresher (younger and less mixed) than that present off Angola and Namibia, and is also fresher than that from the Indian Ocean. The differences in "freshness" of AAIW at three areas in the Benguela is illustrated in Fig. 6. While there is some northward movement evident in the south of mixed South Atlantic and South Indian
AAIW, the greater part of this water mass in the Benguela region (at least in the area adjacent to the continental shelf) evidently moves southwards from the tropical South Atlantic, having reached the area by a somewhat circuitous route. Further offshore, the AAIW flows in a north-westerly direction. AAIW does not upwell to the surface anywhere in the
Benguela.
North Atlantic Deep Water corresponds to the deep salinity maximum (see Fig. 5) and has a salinity typically >34.8
psu, and lies below the AAIW stratum. As its name suggests it is formed in the North Atlantic. It then sinks and spreads southwards. At the equator it comprises a thick layer between 1000m and 3500m of relatively warm (for its depth) and saline water. West of the Benguela continental shelf it flows generally
polewards, becoming diluted
en route south. Volumetrically, NADW is the main water mass present in the South-east Atlantic.
Underlying NADW in the Cape Basin is Antarctic Bottom Water. AABW forms near the edge of the Antarctic continent in the Weddel Sea area, and spreads throughout the Southern Ocean. Unless prevented by topography, it also tends to spread northwards in the South Atlantic, Indian and Pacific Oceans. In the Cape Basin it flows slowly clockwise, moving southwards at depths greater than 4000m west of the Benguela continental shelf. The Walvis Ridge forms a virtually non-penetrable barrier to the northward flow of the
AABW, which as a consequence is not significantly present in the Angola Basin. The effect of the Walvis Ridge on the distribution of AABW is dramatically illustrated in Fig. 6.
Whereas the general movement of deeper water masses viz. AAIW, NADW and AABW in the Southeast Atlantic is
polewards, the flow of the Subtropical Surface Water (STSW) tends to be more closely aligned to the direction of the prevailing wind - at least in the area south of 15°S latitude where the flow is generally in a north-westerly direction with speeds typically in the range 10 - 15 cm/s. with an average of around 17 cm/s. As a consequence of seasonal warming and cooling of surface water, the temperature - salinity characteristics of STSW can be quite variable, and can range from 15° to 23°C and 35.4 psu to 36.0
psu. (see Fig. 5). The surface water present off Angola is mainly of tropical/equatorial origin. Temperatures in excess of 25°C are common, reaching 28° - 29°C in summer and there is usually a very strong and shallow thermocline present. (This and the contrast with the area further south is highlighted in Fig. 6.) Surface salinity in the area is highly variable, ranging from very low values close to the mouths of major rivers such as the Congo, to levels in excess of 36psu further offshore. The influence of the Congo River at the surface can be substantial, and water from this source can be traced as far south as Namibia. (Dr M. E. L. Buys,
pers. comm.) It is generally observed as a thin surface layer, lens-like in places overlying a strongly stratified surface sea water layer.
The principal features of the flow of surface water, and away from the coast also of Thermocline Water, is illustrated schematically in Fig. 7. The broad arrows between 15° and 35°S represent the Benguela Current, which can best be defined as the integrated equatorward flow of the upper layers in the South Atlantic east of the 0° meridian. The circulation has been described in some detail by Stramma and Peterson (1989) and summarised by Shannon and Nelson (1996), and readers are referred to these papers for further particulars. In terms of absolute volume, the total equatorial flow of the Benguela Current, including that of surface, Thermocline and AAIW is thought to be about 15 - 25 Sv (one Sv is 106 m3/s). North of 15°S, which encompasses the Angola system, the main features of the currents within the surface, Thermocline and AAIW layers are (a) a large cyclonic gyre centred around 12°S, 4°E viz. the Angola Dome (b) the Angola Current which flows southwards along the edge of the continental shelf and (c) the Equatorial Undercurrent and South Equatorial Current which feed the Angola Current from the north-west. The oceanography of the Angola system (including the Angola Dome) and the offshore northern Benguela area was well documented by Moroshkin
et al (1970). The existence of the Angola Dome and cyclonic flow around it was confirmed by Gordon and Bosley (1991). The volume transport around the Angola Dome is of the order of 3
Sv. The major dynamics affecting the eastern tropical Atlantic Ocean were summarised Voitureiz and Herbland (1982) and by Picaut (1985).
Of the 15 - 25 Sv equatorward flow in the main Benguela Current (as defined earlier), some 7 Sv is of Indian Ocean origin (Van Ballegooyen
et al 1994). The latter comes from the Agulhas Current which flows southwards and westwards along the east coast of South Africa. The Agulhas Current is the major western boundary current in the Indian Ocean (75
Sv), and is rather analogous to the Gulf Stream and the Brazil Current. The Agulhas tends to follow the edge of the continental shelf and on reaching the Agulhas Bank turns southwards and then eastwards, flowing back into the Indian Ocean. This turning back is termed
"retroflection". Small periodic meanders termed the Natal Pulse, may develop in the main body of the Agulhas Current off Natal and these grow downstream, resulting in the Current becoming unstable and the shedding of large eddies or rings. About six or eight Agulhas rings are shed each year, and these fast spinning rings move slowly (typically 5 -8 cm/s) in a west-north-westerly direction into the South Atlantic, transporting about 7 SV of Indian Ocean water on average. At the surface shallow filaments of Agulhas Current water also may round the Cape of Good Hope, just outside the upwelling area, but the mass of water associated with these filaments is usually fairly small although occasionally the southern Benguela is flooded by warm water of Agulhas origin. The retroflection of the Agulhas Current and the leakage of warm Indian Ocean water into the South Atlantic is well illustrated in Fig. 4, and shown diagrammatically in Fig. 7.
In summary the main features of the surface currents are:
-
Offshore: flow of about 15 - 20 cm/s (i.e. about 1/3 knot in a northwesterly direction over most of the region between 15°S and 35°S (viz. the Benguela Current)
-
Cyclonic circulation around the Angola Dome and periodic intrusion of tropical water from the north and north west into the northern Benguela
-
Leakage of Agulhas Current water into the South Atlantic, mainly via rings and to a lesser extent via shallow filaments
2.5 Shelf circulation
Circulation over the continental shelf and in the oceanic area adjacent to the shelf has been the subject of intensive investigation during the past twenty years, although most of the effort has been focussed on the southern
Benguela. The various observations were summarised by Shannon and Nelson (1996) and again recently by Shillington (1998).
Surface currents over much of the Benguela shelf are largely influenced by the prevailing winds. In the south there is a general convergent flow of surface water from the Agulhas Bank westwards and northwards around the Cape of Good Hope which funnels into a frontal jet west of the Cape Peninsula (discussed later). Typical speeds are in the range of 25 -75 cm/s. Near Cape Columbine (33°S) the surface current divides into an offshore flow and a northward alongshore flow, partially into St Helena Bay. A southward moving current often occurs near the surface close inshore over the entire region, particularly during winter and also periodically during the rest of the year when reversals take place on a time scale of several days. Over the Namibian shelf, the surface currents are generally in a northerly direction, closely aligned to the wind. However, periodic and episodic reversals in the surface currents can occur, the most pronounced and extended reversals occurring during Benguela Nińos (discussed in Section 5). The most prominent circulation feature off the coast of Angola in the southward flowing Angola Current. Whether this current is a permanent or seasonal feature is not clear as most of the more comprehensive investigations appear to have been undertaken during the autumn and winter months. Certainly the existence of the Angola Current was well documented by Moroshkin
et al (1970). It is a coastal current, generally present as a poleward flow over the upper part of the continental slope (i.e. west of the shelf break), detectable between the surface and a depth of about 200m. Literature on the current is sparse, but there is some evidence of seasonality, with the most intense flow occurring during late summer (March), when surface speeds as high as 70cm/s have been reported, and subsurface speeds of up to 88 cm/s
(Dias 1983). The dynamics of the Angola Dome and Angola Current are linked with those of the system of equatorial currents and the Benguela Current and upwelling processes. However, to what extent the Angola Current contributes to the Benguela system off Namibia is uncertain - at the surface and subsurface at least. In this respect Dias (1983) showed that most of the southward flow of the current at a depth of 100m turned between 16°and 17°S latitude to flow westwards just north of the
Angola-Benguela front. At greater depths e.g. 400m, the poleward flow from Angola into the northern Benguela does however seem to be more continuous, and this has been the focus of recent cooperative research between German, Norwegian, Angolan, Namibian and South African scientists.
In the southern Benguela, in particular off the Cape Peninsula, but also in the vicinity of Cape Columbine there exists a strong equatorward flowing jet. First predicted and discovered by Bang and Andrews (1974) the existence of this jet has subsequently been confirmed as a semi-permanent feature of the upwelling system. Current velocities are highest at subsurface depths, being typically in the range 25 - 75 cm/s. The jet is important biologically in that it is known to transport eggs and larvae of various fish species from the spawning grounds on the Agulhas Bank to the nursery areas inshore north of Cape Columbine. To what extent the jet is continuous throughout the
Benguela, or indeed even if it exists in the central and northern Benguela, is not known, although there is a suggestion in the work of Gordon
et al. (1995) that a jet like feature is present in the upper 100m over the mid-shelf near
Lüderitz.
Apart from the shelf-edge jet, the most significant discovery during the past two and a half decades has been of a poleward undercurrent. The idea of a poleward flow in upwelling regions seems to have originated from ideas put forward by Hart and Currie (1960) in their classic text on the Benguela Current viz. as some form of compensation for the water displaced from the inner shelf by upwelling. Nelson (1989) showed that, as in the case of in other upwelling systems, a poleward undercurrent does exist in the
Benguela, but which is much more extensive than that required as compensation for coastal upwelling, stretching from the coast, across the shelf and out into the Cape Basin. Current meters have revealed that subtidal currents on the shelf are dominated by the presence of coastal trapped waves (previously commented on), which have periods of 3 to 8 days. These result in a net polewards flow of about 5km/d or 5.8 cm/s (Nelson 1989). On occasions this southwards-moving current may reach the surface inshore, resulting in episodes of poleward flow at the surface. In St Helena Bay, such events result in episodic flushing of the Bay. Aspects of the poleward current will be discussed further in Section 3.1 which addresses dissolved oxygen.
The shelf-circulation can thus be briefly summarised as follows:
-
Wind driven surface currents over the shelf
-
Poleward subsurface flow over shelf and in deeper water adjacent to shelf throughout the region (4-5 km./d)
-
Poleward flowing Angola Current in extreme northern part of the Benguela region i.e. along coast of Angola
-
Jets associated with the upwelling system and located near the edge of the continental shelf in the southern Benguela
-
Coastal trapped waves are characteristic of the shelf area throughout the Benguela (periods typically 3-8 days).
2.6 System boundaries, fronts and filaments
Whereas a thermocline refers to a layer where there is a rapid change in temperature with depth, a halocline and a pycnaocline are vertical discontinuities of salinity and density respectively. Fronts refer to areas where there is a sharp change in temperature, salinity (or some other parameter such as
colour) horizontally. Reference back to Fig 3 which illustrated a simplified concept of upwelling, it can be seen that where the thermocline intersects with the sea surface, the vertical discontinuity is translated into a horizontal discontinuity viz. a front. Physical boundaries of, and within, the Benguela are usually associated with fronts of one form or another, and these fronts tend to form barriers to horizontal movement of water and small particles such as plankton. They may be less important as barriers for highly motile animals such as fish and marine mammals.
Northern boundary
The physical northern boundary of coastal upwelling is marked by the Angola-Benguela frontal zone. The temperature and salinity front (series of fronts) is a permanent feature at the surface, identifiable to a depth of at least 200m, and is maintained throughout the year within a narrow band of latitudes, characteristically between 14°S and 17°S (i.e. close to the Angola-Namibia border). The front generally has a west-to-east orientation, and appears to be maintained by a combination of factors, including bathymetry, coastal orientation, stratification, wind stress and opposing flows of the Benguela and Angola Currents. The southwards migration of the front is most pronounced during late summer when longshore winds in the northern Benguela are weaker and upwelling is reduced. The situation in northern Namibia/southern Angola is in some respects analogous to the seasonal cycle off Peru in the Pacific during some years, when anomalously warm water appears
(El Nińo). There is a South Atlantic equivalent of the Pacific El Nińo and it manifests itself as an episodic extreme warming in the tropical eastern Atlantic and the movement of tropical water southwards and eastwards along the Namibian coast. These
Benguela Nińos (Shannon et al. 1986) occur on average every ten years and are not necessarily in phase with the
El Nińo- Southern Oscillation (ENSO) - although some links with the latter are evident.
Benguela Nińos occurred in 1934, 1949, 1963, 1984 and 1995 and probably also in 1910, in the mid-1920s and in 1972-1974. Although not as frequent or intense as
El Nińos, like the Pacific counterpart, their impact on the ecosystem and harvested resources in the northern Benguela is enormous.
The characteristic location of the Angola-Benguela frontal zone is shown in Fig. 7. This is a much simplified diagram, as the "front" is often a combination of fronts with convoluted characteristics. (For example see Fig. 8 - which illustrates the surface expression of the
Angola-Benguela frontal zone in April 1997).
While the Angola-Benguela Front (more correctly a series of fronts) comprises the northern extent of the main coastal upwelling zone, upwelling can occur seasonally along the entire coast of Angola. There are, in any event, strong linkages between the behaviour of the
Angola-Benguela front (and the oceanography of the area to the south of it) and processes occurring off Angola, especially the Angola Dome and the Angola Current. Unless these are considered as an integral part of the
BCLME, it will not be feasible to evolve a sustainable integrated management approach for the
Benguela. Moreover, there is a well-defined front at about 5°S, viz the Angola Front which is apparent at sub-surface depths. It is this front which is the true boundary between the Benguela part of the South Atlantic and the tropical/equatorial Gulf of Guinea system. A northern boundary at 5°S would thus encompass the Angola Dome, the coastal Angola Current and the area in which the main oxygen minimum forms and the full extent of the upwelling system in the South-east Atlantic. A pragmatic northern boundary is thus at 5°S latitude (see Fig 7) which is in the vicinity of the northern boundary of Angola
(Cabinda) and the southern extent of the Gulf of Guinea Large Marine Ecosystem (GOGLME).
Southern boundary
The southern boundary of the Benguela system can be considered as the Agulhas retroflection area, typically between 36° and 37°S. Like the northern boundary, this warm southern boundary pulsates on a spectrum of time and space scales, and about 10% of the warm tropical Agulhas Current "leaks" into the South Atlantic. As previously explained most of this leakage is in the form of rings (eddies) which are shed from the Agulhas Current as it retroflects. The main trajectory of these shed rings is west-north-west, although departures from this have been recorded, and there exists a well documented case of ring interacting with the Benguela upwelling system and drawing upwelled water off the shelf in the form of a large curved upwelling filament. In addition to rings, there is a small regular leakage of filaments of Agulhas water around the Cape of Good Hope just west of the edge of the shelf and thermal front (which is associated with the upwelling). On occasions there are substantial intrusions of Agulhas Current water into the southern Benguela, of which the best documented case occurred in 1986 and coincided with that year being among the warmest this century in the South-east Atlantic Ocean. Another large intrusion occurred during the summer of 1997/8. Like its counterpart in the north, these Agulhas intrusions appear to affect the living resources in the southern Benguela.
The location of the southern boundary is shown schematically in Fig.7, while the typical extreme complexity of this boundary and retroflection of the Agulhas Current and "leakage" is illustrated in Fig. 4.
Western boundary
The western (offshore) boundary of the Benguela is fairly open ended, but is generally taken as approximately the 0° meridian. As such the Benguela
sensu lato includes the coastal upwelling area, the longshore fronts (see below) and the eastern portion of the South Atlantic gyre. By
definition then the Benguela Current comprises the total area of equatorward flow in the upper part of the South-east Atlantic Ocean.
Longshore fronts
There exists over much of the area between Cape Frio (18°S) and Cape Point (34°S) a well-developed longshore temperature front or fronts, which extends seasonally (in summer) eastwards around the Cape of Good Hope. The oceanic thermal front approximates to the seaward boundary of the general area influenced by coastal upwelling. South of Lüderitz a single front is usually well defined, and although spatially and temporally variable, coincides approximately with the run of the shelf break (edge of the continental shelf). Further north the surface manifestation of the front is more diffuse and multiple fronts are evident on occasions. The meandering nature of the front is evident in satellite imagery (see Figs 4 and 8). Upwelling
filaments which have a life span of days to several weeks and which are generally orientated perpendicular to the coast cause the front to become highly convoluted. Opinion as to whether these filaments are randomly distributed or site specific are divided, although most recent evidence points to the latter (which can be explained in terms of bathymetry and system dynamics).
3. CHEMISTRY AND RELATED PROCESSES
Prior to 1925 very little was known about the chemistry of the South-east Atlantic Ocean and adjacent areas, and it was not until data collected by the German
Meteor Expedition of 1925 - 1927 were analysed and interpreted that a picture of the large-scale distribution of elements of importance such as oxygen, phosphorous and silica began to emerge. In the Benguela region, although the role of upwelling in the supply of nutrients (the dissolved fertilisers in the sea) was appreciated as early as the 1930s, it was not until the publication of the definitive work of Hart and Currie (1960) that the nutrient chemistry was placed in a proper physical and biological perspective. Subsequent investigations by
inter alia authors such as De Dekker (1970), Calvert and Price (1971) and Andrews and Hutchings (1980) resulted in a greatly improved understanding of the chemical-biological processes of importance in the Benguela. The available information on the Benguela chemistry and related processes was reviewed by Chapman and Shannon (1985) while recently Bailey and Rogers (1997) have provided a useful overview of chemical oceanography within the context of marine geoscience in southern Africa.
Before discussing the essential features of processes associated with the chemistry of the Benguela system it is perhaps appropriate to explain some basic concepts so that readers who are not oceanographers or chemists will more easily be able to follow the subsequent discussions. Viewed very simply, macro-nutrients such as ammonia, nitrate, phosphate and silicate are present in sea water at low but significant concentrations. Near the sea surface these nutrients are consumed by microscopic plants (phytoplankton). Photosynthesis takes place, and the phytoplankton multiplies or blooms, absorbing carbon dioxide and releasing oxygen into the water. The phytoplankton is then either consumed by small planktonic animals (zooplankton) and some fish species or otherwise sinks slowly to the bottom. In this process bacteria play an important role, and as the phytoplankton and also faeces and organic material from zooplankton and higher consumers sinks, these decay. During the decay, oxygen dissolved in the deeper sea water layers and sediments is consumed and nutrients are released back into the water column. The overall process results in a lowering of nutrient concentrations in the surface layers and an increase at depth, with the opposite situation applying to dissolved oxygen. Within the context of a system such as the Benguela, it is the upwelling that is thus so important in bringing the nutrient-rich deeper water to the surface where photosynthesis can occur, and this explains why eastern boundary currents are so rich biologically. In the Benguela there is an imbalance between phytoplankton and zooplankton and fish production, and much of the decaying phytoplankton is deposited on the seabed, forming the organically rich diatomaceous sediments which are characteristic of the larger part of the Benguela shelf. In turn, processes which take place at the interface between the sediments and the overlying water and in the water in the sediments (interstitial water) are chemically also very important, in particular for the "regeneration" of nutrients.
Although what has been described is perhaps somewhat of an oversimplification of the complex physical-chemical-biological interactions which occur, it does help to explain the cycle.
3.1 Dissolved oxygen
One of the major features of the Benguela region is the occurrence of large areas where very low concentrations of dissolved oxygen are found. Thermocline Water (Central Water) in the South-east Atlantic Ocean, which is the upwelling source water, commonly contains between 4.8 and 5.2 ml/l dissolved oxygen and is about 80 - 85% saturated. In contrast, the shelf waters in the Benguela system frequently contain much lower concentrations, and on occasions at sites such as Walvis Bay the sea can become anoxic at times, particularly at depth, while near the surface during phytoplankton blooms photosynthesis can result in the surface layer becoming supersaturated with oxygen. The occurrence of low-oxygen water is not only important in terms of the chemistry, but plays a key role in controlling the distribution and abundance of several marine species - not only bottom-dwelling (benthos) like rock lobster, but also fish such as hake. To avoid confusion, in the following paragraphs, we shall refer to water having an oxygen content of less than 2 ml/l as "oxygen-deficient" and above 2 but less than 5 ml/l as "oxygen-depleted".
The first comprehensive account of the large scale distribution of dissolved oxygen in the South Atlantic was provided by Wattenberg (1938), and was based on measurements made during the expedition of the
Meteor a decade earlier. His work showed the existence of a wedge-shaped tongue of oxygen-deficient water with its core lying at a depth of 300 - 400m, and extending from its base between the equator and 20°S at the African continent, across the South Atlantic. Concentrations lower than 0.5ml/l were recorded in the Benguela region near 15°S. The permanence of this major low oxygen feature in the South Atlantic has been confirmed by all subsequent large-scale investigations.
Hart and Currie (1960) and others have demonstrated the existence of an oxygen-deficient layer overlying the continental shelf north of Walvis Bay, and it is clear that oxygen-depleted subsurface water is a characteristic feature of much of the northern and central Benguela shelf. Subsequent studies have shown that low oxygen conditions can exist at times on the shelf further south, for example near the Orange River, in St Helena Bay and even at some sites on the Agulhas Bank. Hart and Currie (1960) suggested that the oxygen-depleted shelf water might be transported southwards from the tropical South-east Atlantic along the edge of the shelf in a deep compensation current, a view supported by De Dekker (1970) and Andrews and Hutchings (1980) who put forward convincing arguments in favour of this, and suggested that the oxygen-deficient water which occurs as far south as the Cape Peninsula at times, originates from Namibia.
The most comprehensive study of dissolved oxygen off Angola and Namibia was that undertaken by Bubnov (1972), and he suggested that the main oxygen minimum in the South Atlantic forms in a broad area off Angola, and is reinforced by processes associated with the Angola Dome. This is shown schematically in Fig. 9. Bubnov, however, calculated that the level of primary production over the Namibian shelf was sufficient to provide the concentrations of oxygen observed in the water column there in the absence of any advection (horizontal movement) from the north, a view supported subsequently by a number of authors. Interestingly, Poole and Tomczak (1999) show that the pseudo age of this oxygen poor water off Angola is 50 years or greater, in contrast to the younger more recently ventilated water south of 30°S which is less than 10 years "old".
Thus, while southward advection of oxygen-deficient and oxygen-depleted water from the Angola Basin via a poleward undercurrent may be an important mechanism controlling the distribution of low oxygen water on and adjacent to the shelf, local processes over the Namibian shelf are probably more important
on average as determinants of oxygen dynamics in the Benguela region per
se. This view is reinforced by the properties and distribution of the main oxygen minimum layer off Angola which are different to those of the oxygen-depleted/deficient shelf waters off Namibia. Indeed two distinct maxima cores have been observed on occasions, one over the shelf (locally produced) and a deeper one (probably part of the main minimum layer) west of the shelf break. It does seem, however, that the high primary production off Namibia is an important contributor to the main oxygen minimum layer, the latter at times spilling onto the shelf and reinforcing the depletion processes in Benguela shelf waters. There is also evidence that there is a slow southward advection of oxygen-deficient water throughout the Benguela at least as far south as the Cape Peninsula with lowest oxygen concentrations occurring during late summer/autumn in the southern Benguela. The fact is that oxygen-deficient water dynamics which plays a pivotal role in the ecosystem are not well understood. What is known is that substantial interannual variability in the oxygen concentrations does occur (discussed later) and that this is important for fisheries.
A conceptual model of areas where the low oxygen water forms and its movement is given in Fig. 9. Readers are referred to Chapman and Shannon (1987) and Bailey and Rogers (1997) for further information.
3.2 Nutrients
The general features of the distribution of nutrients in the Benguela resemble closely those of other upwelling regions. The upwelling water is enriched in nutrients relative to the surface layers and during active upwelling this water reaches the euphotic zone (the biologically productive surface layer which sunlight penetrates) near the shore. Following the establishment of the thermocline, phytoplankton production consumes nutrients in the upper layers, leaving them much depleted, while nutrient re-enrichment occurs below the thermocline as the phytoplankton decay. Chapman and Shannon (1985) pointed out the difficulty in discussing the nutrient status of the whole Benguela because the chemistry is very much site specific, and it is therefore not easy to generalise.
The shelf waters of the Benguela are, however, characterised by elevated concentrations of nutrients in comparison with those in the surface mixed layer of the adjacent oceanic waters, and also in comparison with concentrations in source waters. For example, South Atlantic Thermocline Water contains about 0.8 - 1.5 mM (micro-moles) phosphate, but shelf waters have phosphate concentrations typically between 1.5 and 2.5 mM, with values as high as 8mM having been recorded off Namibia. Local regeneration processes are important throughout the Benguela, but particularly off Namibia. In comparison with the eastern boundary of the Pacific, source waters in the Benguela have lower levels of inorganic nutrients, and consequently a lower potential for new production. (The term "new production" is commonly used by marine chemists and biological oceanographers and can be viewed simply as that based on outside sources of "fertilisers" such as nitrates, unlike " regenerated production" which results from the locally produced waste material - ammonia and urea based.). Typical concentrations of macronutrients in the Benguela system are summarised in Table 1.
Table 1: Nutrient concentrations (mM) in (a) offshore upwelling (b) shelf and (c) oceanic surface waters in three areas of the Benguela, based on published work.
|
Area |
|
Nitrate |
Phosphate |
Silicate |
|
Cape
Peninsula |
(a) |
20 |
1.5 |
16 |
|
|
(b) |
23 |
1.5 |
19 |
|
|
(c) |
<1 |
0.5 |
5 |
|
St Helena Bay - Orange
River |
(a) |
25 |
2 |
10-20 |
|
|
(b) |
25 |
2.5-3 |
20-40 |
|
|
(c) |
<1 |
0.5-1 |
8 |
|
Namibia |
(a) |
15-25 |
1.5-2.5 |
5-20 |
|
|
(b) |
10-30 |
2-3 |
20-50 |
|
|
(c) |
<5 |
<2 |
<1 |
One generalisation which can be made is that the importance of nutrient regeneration in the supply processes increases northwards in the Benguela system.
Whereas Thermocline (Central) Water of South Atlantic origin contains fairly similar concentrations of nitrate and phosphate as South Indian Ocean Thermocline Water, the silicate content of the latter is only about half of the former. This is significant for the nutrient chemistry of the Agulhas Bank region, which can according to Lutjeharms
et al. (1996) be divided into distinct nutrient "provinces," the eastern off shore part of the Bank being dominated by nutrient-poor Subtropical Surface Water and the western and inshore areas influenced by nutrient-rich South Atlantic Thermocline Water. Although upwelling occurs inshore in the lee of prominent capes, the Agulhas Bank system is strongly stratified in summer, particularly so in the eastern part. As a consequence nutrient concentrations tend to be higher in winter over the Bank as a whole in winter when the shallow seasonal thermocline breaks down. Readers requiring further information are referred to an excellent overview of the nutrient dynamics of the Agulhas Bank in Bailey and Rogers (1997).
Published work suggests that silicate is the limiting nutrient at times in the northern Benguela, while nitrate is on occasions limiting in the southern part of the system. This is perhaps counter intuitive when it is considered that the Namibian shelf sediments are rich in biogenic silica and Agulhas Current/Bank water (which leaks into the southern Benguela) relatively poor in silica. Considerable advances have been made during the past decade with respect to the assimilation of ammonia and nitrate by phytoplankton using stable-isotope nitrogen fifteen (N 15) incubation techniques by authors such as Probyn
et al. (1990), Probyn (1992) and Waldron and Probyn (1992).
A significant contribution to the understanding of nutrient dynamics and production was made during the second phase of the Benguela Ecology Programme (1987-1992) with the recognition of the importance of the microbial loop and the application of N 15 labelling techniques to distinguish between newly incorporated nitrogen (NO3-N or N2) and metabolically recycled nitrogen (NH4-N or dissolved organic-N, i.e. urea).
Using satellite-derived sea surface temperature imagery, Waldron and Probyn (1992) estimated the potential new production in the Benguela system, and the first of these authors subsequently went on to calculate new production during the 1980s using sea level as a proxy for upwelling. Following on from work on carbon pathways in the southern Benguela upwelling system (Waldron
et al. 1998), Dr Waldron has kindly generated a nitrate-nitrogen driven pathway for the Benguela as a whole for the purpose of this overview. This is given in Fig. 10 and illustrates simply and dramatically the import and export fluxes in shelf waters, in particular the nitrate-driven new production and the role of the shelf sediments as a sink. This will be discussed further in the section dealing with the foodweb and carbon budget.
3.3 Sulphur
Perhaps one of the most obvious features of the marine chemistry of the northern Benguela is the odour of hydrogen sulphide gas which is associated with "sulphur eruptions". These periodic eruptions are common in the general vicinity of Walvis Bay, usually during late summer when upwelling is at a minimum i. e. under quiescent conditions.
Sulphide formation results when anaerobic biological breakdown of organic substances and bacterial reduction of sulphate which is present in sea water and in the interstitial water in the marine sediments occur. As biological reduction of sulphate is an anoxic process, sulphide formation can occur where there is above average oxygen consumption or poor circulation - circumstances which may be associated with the decay following a major bloom of phytoplankton in an embayment or even along an open coast under quiescent conditions. These suboxic or anoxic conditions are common in the northern Benguela shelf waters and underlying organically rich sediments, and this provides suitable environmental conditions for the formation of sulphides by sulphate reducing and anaerobic bacteria.
As hydrogen sulphide which may be formed in the process can be extremely toxic, even at very low concentrations, mass mortalities of marine organisms are often associated with the "sulphur eruptions"' compounding the effect of the already depleted oxygen content of the sea water. Records of fish kills in the Benguela resulting from sulphide production go back at least as far as 1928, but the occurrence of sulphide was not positively identified until later (Copenhagen 1934). Sulphurous fumes are often present in the atmosphere at coastal sites in central Namibia and may penetrate 60km or more inland. Their corrosive effect on iron and steel and tarnishing of paintwork, brass and silver is visibly evident in Walvis Bay and Swakopmund. During the sulphide events, mud islands may appear and disappear, the smell of rotten eggs pervades the air, and the sea takes on an appearance of boiling - hence the term "sulphur eruptions". The most recent widescale sulphur eruption occurred in the Walvis Bay/Swakopmund area during March/April 1998 and was characterised by a strong odour of hydrogen-sulphide. The sea along the coast turned a milky-turquoise colour for as far as the eye could see.
The main area within which the sulphur eruptions occur is the so-called "azoic zone", and free sulphur can be present in sediments from this area. Even 100km further south, concentrations of sulphide as high as 65mg/l may occur in interstitial water in sediments (Bailey 1979). Hydrogen sulphide concentrations of about 1ml/l have been measured in the northern Benguela. In the southern Benguela, eg. St Helena Bay, even though sediments may smell of sulphide, there are very few published measurements of free hydrogen sulphide. However, during the autumns of 1994 and 1998 a strong odour of hydrogen sulphide persisted for about a week in the atmosphere around Cape Town, as a consequence of decaying phytoplankton blooms in the St Helena Bay area, suggesting that sulphide events may be more common in the southern Benguela than hitherto appreciated.
In spite of the occurrence of sulphide in Benguela sediments and shelf waters, surprising little research has been conducted on the subject. Apart from the relevance of such studies to investigation and prediction of mass mortalities of marine life, public awareness and interest is high. Moreover, what is perhaps not well known, but may be extremely important in terms of industrial development along parts of the west coast, relates to the solubilization of heavy metals. For example mercury sulphide which is insoluble in normal oxygenated (oxic) water and sediments and is generally regarded as environmentally harmless, can be transformed to polysulphides of mercury in the presence of low concentrations of sulphide, and goes into solution - thereby becoming a serious environmental hazard.
3.4 Other aspects of marine chemistry
In their comprehensive review of the chemical oceanography of the Benguela system, Chapman and Shannon (1985) also discussed other aspects of the redox chemistry, including that of iodine and biomine, in addition to nitrate/nitrite and sulphate/sulphide as couples. (An important reference work on the subject is Price and Calvert 1977 which complements an earlier publication by these authors viz. Calvert and Price 1971). Chapman and Shannon also reviewed published work on the minor elements, which in contrast to the oxygen and nutrient distributions and dynamics have received very little attention in the Benguela. Minor elements include
inter alia the alkali elements (potassium, rubidium and lithium), barium, and heavy metals. Many of the pioneering measurements of metals such as copper, iron and manganese, which are trace nutrients, were made by Orren (1969, 1971). As part of a South African marine pollution study, these and trace metals such as cadmium, nickel, lead and zinc were measured in water and sediments along the coast during the 1970s and 1980s eg. Eagle
et al. (1982). Their results were summarised by Shannon and Chapman (1985) as was the work of authors such as Chester and Stoner (1975) on the concentrations of metals in dust and surface water particles collected in the Benguela region. Excluding urban environments, most of the measurements on metals in sediments have been from the organically rich Namibian shelf region, and it would appear from the available data that the concentrations of metals in these sediments are higher than observed in many other marine sediments.
Man-made chemicals such as chloro-fluoro-carbons ("CFCs") have been detected in the deeper water masses present in the South-east Atlantic. CFCs such as CFM-11 and CFM-12 are useful tracers of water mass age and movement, and in the southern African context of the exchange of waters between the South-east Atlantic, North Atlantic and Indian Oceans.What the results also show is that even the deep bottom waters in the Southern Hemisphere have potential to be contaminated by activities of industrial Northern Hemisphere countries and can no longer be considered as pristine.
In conclusion it must be noted that chemical oceanography has for the past two decades been very much a "Cinderella discipline" in southern African countries, and in spite of the importance of chemical processes in regulating the biology of the Benguela ecosystem, this has apparently not been adequately recognised by the fisheries and environmental agencies.
4. PLANKTON AND THE FOODWEB
The literal translation of the word "plankton" is "wanderer", and the term applies to a spectrum small neutrally buoyant ("free floating") organisms in the sea which have little or no power of locomotion, and which drift with the currents. These small microscopic plants and animals provide the primary food sources for a host of marine species and constitute the early building blocks in the marine food chain or, more correctly, the foodweb. The microscopic unicellular plants which occur in surface layers of the ocean are known collectively as phytoplankton, while their animal counterparts are referred to as zooplankton. Although the most common components of zooplankton are only a few millimeters in size, zooplankton includes much larger floating animals such as jellyfish. Useful reviews of plankton in the Benguela system are those of Shannon and Pillar (1986) - now a bit dated - and Hutchings and Field (1997). Perhaps the most readable introduction to the zooplankton of the Benguela Current region is Gibbons (1999).
4.1 Phytoplankton and primary production
Like all plants, phytoplankton require sunlight in order to photosynthesise, and accordingly most of the organic productivity associated with these floating algae takes place near to the sea surface. Phytoplankton can for all practical purposes be divided into two categories viz. diatoms, which have no power of self propulsion and which have an outer skeleton of silica, and those which have small hairs or flagella which enable some weak motion viz. flagellates. The latter are generally more fragile than diatoms and are associated with less-turbulent and more stratified waters. Diatoms are characteristic of turbulent, nutrient-rich upwelled water. In upwelling systems the biomass of diatoms is generally much higher than the biomass of flagellates.
The Benguela is generally regarded as a diatom-dominated system. This perception is to some extent an artifact of past sampling, which has tended to miss the very small cells or nanoplankton. (The productivity of nanoplankton is regulated by
regenerated nitrogen - see Section 3.2 - and historically nanoplankton have been undersampled in the Benguela). Both the northern and southern Benguela share many similar species assemblages, with
Chaetoceros, Nitzschia, Thalassiosira, Rhizosolenia being endemic throughout the region. There are, however, essential differences between the north and the south, some of which are linked to the atmosphere/ocean dynamics (e.g. nutrient supply, turbulence and stratification). The diatom
Delphineis karstenii (Fragilaria karstenii) is restricted to the north, while
Skeletonema costatum is found predominantly in the southern Benguela, evidently having been most abundant in the early-mid 1960s and mid 1980s (both warm periods in the system). The large cell
Coscinodicus spp. commonly occurs in areas of high turbulence. Over the Agulhas bank, the species assemblages are more cosmoplitan than along the west coast. Microflagellates are common in the central area of the northern Benguela e.g.
Gymnodinium and Peridinium spp.
Phytoplankton abundance both from net and bottle sampling and chlorophyll a measurements highlights the dichotomy between the northern and southern parts of the system, with low values around 27-28°S (the base of the Lüderitz upwelling cell), and high values downstream of the cell, and also further south associated with downstream areas of the other upwelling cells (Namaqua, Columbine, Cape Peninsula). Off Namibia during periods of active upwelling, highest concentrations of phytoplankton occur offshore (50km), and during quiescent periods in a narrow band close to the coast. A characteristic difference between the northern and southern Benguela is that concentrations of chlorophyll a are generally higher off Namibia than off South Africa, being more uniformly distributed in the former with less well defined chlorophyll fronts at the oceanic boundary.
The advent of ocean colour satellite imagery in the late 1970s provided new insights into the distribution and large scale dynamics of phytoplankton blooms. The Coastal Zone Colour Scanner (CZCS) on the Nimbus-7 satellite in fact resulted in a quantum leap in knowledge and understanding of the global distribution of phytoplankton, and southern African oceanographers played an important role in the early validation of CZCS data and application of satellite-derived ocean colour imagery in the study of upwelling ecosystems and fisheries. After about ten years (which is a very long time for a satellite sensor to function) the CZCS eventually failed, and it wasn't until very recently that a new and much more sophisticated generation of colour satellites were launched and became operational. Of these, the Sea-viewing Wide-imaging Field-of-View Sensor (SeaWiFS) launched in August 1997 is the best-known and most user-friendly. Fig. 11 is a snapshot of the distribution of plant pigments in the Benguela between 16° and 32° S from SeaWiFS.
In the southern Benguela chlorophyll is determined by wind cycles and displays significant seasonality. Maximum concentrations tend to occur 20-80km offshore. (Blooms following periods of active upwelling can extend 100km or more offshore). Chlorophyll a concentrations in recently upwelled water, maturing upwelled water and aged upwelled water are about 1, 1-20 and 5-30 mg/m3 respectively. Over the Agulhas Bank phytoplankton production is largely controlled by thermocline/nutricline dynamics and the area is characterised by a deep chlorophyll maximum layer, at a depth of about 40m.
The total primary production in the Benguela system is approximately the same as that in the Peruvian system, but substantially greater than off California. Brown
et al. (1991) provided an authoritative review of phytoplankton primary production and biomass in the Benguela. Average values are given in Table 2.
Table 2. Average values of phytoplankton biomass and production.
|
|
Phytoplankton Biomass |
Primary production
(C14 uptake) |
|
Tons C |
g C/m2/d |
tons C/y |
|
Northern Benguela (15°S-28°S) |
2.6x106 |
1.2 |
77x106 |
|
Southern Benguela (28°S-34°S) |
0.7x106 |
2.0 |
76x106 |
|
South-west Coast (34°S-30°E) |
0.5x106 |
1.9 |
79x106 |
Bacterial biomass and production estimates range from a conservative 2-7% and 3-5% of that of phytoplankton to 8-27% and 26-44% respectively in the least conservative case.
Substantial advances have been made with respect to the dynamics of phytoplankton-blooms and plankton-ecology in general in the southern Benguela by drogue studies (following patches of freshly upwelled water as it ages) and anchor station experiments. (The latter were also done in the northern Benguela.) During the past decade a good understanding of community structure and variability has been gained in the southern Benguela which has showed systematic trends in dominant patterns of diatoms, dinoflagellates and microflagellates in relation to upwelling, pulsing, turbulence and stratification. The importance of phytoplankton seed populations in determining the composition and time required for bloom development has been established and insights have been gained into maintenance and life-survival strategies. In this respect, diatoms tend to form spores which sink rapidly, and this enables them to remain in the nearshore and nutrient-rich environments closer to upwelling centres (Pitcher 1990). Studies on rate dynamics have enabled the quantification of the phytoplankton loss process to be made. The knowledge gained in the south, if applied in the north, should facilitate a quantum jump in the understanding of the dynamics of the Benguela - at least in respect of plankton processes.
4.2 Red tides and harmful algal blooms.
Outbreaks of red tide occur both in the northern and southern Benguela. They tend to be observed most frequently close inshore, where their visual and at times harmful effects are most apparent. Red tides are most frequent during quiescent conditions which follow upwelling, or during periods of light onshore winds and downwelling which commonly occur in the southern Benguela during times of Pacific
El Nińo events. The red tide causing organisms in the Benguela are generally dinoflagellates and sometimes ciliates, and contrary to popular belief, most of the red tide species are non-toxic (Horstman 1981).
That red tides occur regularly in the Benguela is not unexpected. Diatoms, which dominate the phytoplankton in the Benguela, have high nutrient requirements and are adapted to turbulent conditions. The diatom blooms which are associated with upwelling events quickly strip the nutrients from the upwelled water, and as the water column stabilises and stratifies, this provides a suitable environment for dinoflagellates, which being tolerant of low nutrient conditions, and favouring stable high light conditions, can then outcompete the diatoms and "bloom". The dynamics of red tide blooms in the Benguela have been studied in some detail, in particular in the south, during the past two decades by Dr G. Pitcher and his coworkers. In a recent article Pitcher and Boyd (1998) have described from an examination of the distribution of dinoflagellates across the continental shelf and of currents the physical mechanisms responsible for red tide outbreaks, and for the maintenance of motile populations within the system.
The most common red tide organism in the southern Benguela is Noctiluca scintilans, a non-toxic dinoflagellate. It also occurs off Namibia. This organism has been associated with fish mortalities, however, not through any toxin, but by depleting the dissolved oxygen in the water during major blooms, and also evidently by clogging the gills of fish.
Noctiluca blooms have a characteristic bright orange colour - almost luminous (see Fig. 12). The most common red tide species off Namibia appears to be
Heterocapsa triquentra, and like species of Gymnodinium, Gongaulex and
Scrippsiella, it has been linked to mortalities in fish. Most reported mortalities of marine life in the Benguela which have been associated with harmful algal blooms (HABs), in particular mortalities of sand mussels and benthos have been due to a few species of
Gongaulex and to Mesodinium rubrum. The latter species is also known to occur off Angola. Red tide outbreaks off Angola were first reported in the scientific literature of the 1940s. A major bloom of
Prorocentrum balticum occurred between Namibe and Luanda in August-September 1951, and was associated with high fish mortality (Silva, 1953) - see also a comprehensive account of red water on the coast by Paredes (1962). Pitcher (1999) recently reviewed the pertinent information on HABs in the Benguela system. His report is an excellent overview of the subject and introduction to the published literature.
Whether or not the frequency of red tides and HABs in the Benguela is increasing, is not clear. What is known is that there are interannual fluctuatations in occurrences, probably associated with changes in weather patterns. Moreover, in the late 1980s, "green tides" occurred in the area east of Cape Point, in particular in False Bay. These outbreaks caused large mortalities of sedentary organisms such as abalone and also led to respiratory problems in humans swimming in, or in close contact with, affected areas of sea water. The species responsible was
Gymnodinium nagasakiensis and is though to have been imported via ballast water discharges. However, relatively little is known about the extent of exotic species introduced into the system from such discharges.
4.3 Zooplankton and secondary plankton
Zooplankton in the Benguela ecosystem is dominated by small crustaceans (tiny shrimp-like animals), the most important groups being copepods and euphausiids. Of these copepods are numerically the most abundant and diverse group. Species diversity is highest near the warm water boundaries of the ecosystem i.e. in the vicinity of the confluence between the Angola and Benguela Currents, west of the oceanic front and shelf break, and in the extreme south over the Agulhas Bank and adjacent Agulhas retroflection area. Over the shelf, within the main upwelling system copepod diversity is lower, and biomass higher, being dominated by a mixture of small (Paracalanus, Ctenocalanus, Oithona, Clausocalanus), medium (Centropages, Metridia) and large (Calanoides, Rhincalanus) copepods. Among the most common species are
Centropages brachiatus, Calanoides carinatus and Metridia lucens. On the Agulhas Bank the zooplankton biomass is dominated by a single species, the large copepod
Calanus agulhensis which has a centre of distribution in the central-eastern part of the Bank - over the endemic ridge of cool water. Copepods play an important role in the trophic functioning of the Benguela ecosystem. They are the principal food of anchovies and as a consequence they are the most studied zooplankton group - in the southern Benguela at least.
There are more than 40 species of euphausiids in the Benguela ecosystem. Of these
Euphausia lucens is the dominant euphasiid in the southern Benguela and Nyctiphanes capensis in the north. Except in the central Benguela i.e. near Lüderitz, these two species generally do not occur together. The latter species also occurs, however, east of Cape Agulhas. Overall abundance of euphasiids appears to decrease towards the boundaries of the Benguela ecosystem. Studies on the life history of euphausiids in relation to the physical environment have led to an improved understanding of the role of euphausiids in the ecosystem and also to a better understanding of the dynamics of the various fronts and associated upwelling and sinking processes. Studies of the vertical distribution and daily movement of euphausiids in the Benguela have shown that the younger stages to occur near the surface and migrate little, while older stages occur deeper and display significant migration. Euphausiids are important prey items for anchovy and hakes, and conversely
E.lucens is capable of capturing and consuming small fish larvae.
Thaliaceans (salps and doliolids - gelatinous zooplankton) are common throughout the Benguela. They are often indicators of intrusions of warm water, particularly in the southern Benguela. Analyses of gut content suggest that thaliaceans feed mainly on phytoplankton in inshore waters and on zooplankton offshore. The impact of thaliaceans on zooplankton and ichthyoplankton (fish eggs and larvae) has not been quantified, but it could be significant at times. Likewise the abundance and impact of other gelatinous zooplankton, which is periodically abundant in the Benguela, in particular off Namibia ("jelly invasions"), has not been quantified.
In the northern Benguela peak abundances of zooplankton appear to coincide with periods of maximum phytoplankton abundance viz. November - December and March - May, the former following the main upwelling season and the latter during moderate upwelling when summer stratification weakens. In both cases the spatial distribution of zooplankton differs from that of phytoplankton in that zooplankton tend to be more abundant offshore of the (coastal) phytoplankton maxima (beltlike distribution parallel with the coastline). This is probably an oversimplification. However, as there is a general lack of proper quantitative estimates of zooplankton abundance and production in the northern Benguela, coupled with the fishery-centred sampling bias.
In contrast, in the southern Benguela there are relatively good estimates of zooplankton distribution, abundance and production. Zooplankton standing stock estimates in the upwelling area off the Cape Peninsula display distinct seasonality, associated with the upwelling cycle, with a winter minimum and a summer maximum. Superimposed on the seasonal cycle is substantial shorter period variability (determined by upwelling pulsing, the dynamics of the phytoplankton blooms and the life histories of the various zooplankton groups). The series of drogue studies in the 1980s which followed an upwelling pulse and tracked the processes as the freshly upwelled water parcels aged, have contributed substantially to the understanding of plankton dynamics. In the southern Benguela the best estimates of zooplankton production suggests that it is of the order of 80gC/m2/y.
Hutchings et al. (1995) have reviewed inter alia zooplankton grazing in upwelling systems and highlighted a number of generally applicable principles which follow: Although copepods and euphausiids have rapid responses to increased food in terms of egg production, their response in terms of growth of juvenile stages to adulthood are much slower. Behavioural adaptations promote maintenance in the upwelling circulation. Juvenile stages remain near the surface; older stages migrate more extensively and are advected back into inshore water again, allowing grazers to prolong contact with phytoplankton blooms. Wind reversals/calms which allow phytoplankton to return shorewards or be entrained in eddies as they develop may increase the phasing effiencing between phytoplankton and zooplankton. Because of their limited mobility power, zooplankters need to adapt behavioural responses to maximise contact with food items and also minimise predation mortality. The recirculation of zooplankton may be favoured by undercurrents, eddy structures - such as exist near upwelling centres - and inshore counter currents. Like upwelling systems in general, in the Benguela zooplankton biomass maxima tend to exist downstream from upwelling centres and it is these areas which are preferred habitats of developing juveniles of fish species such as anchovy and sardine. Examples of these areas are St Helena Bay, Orange River bight, near Walvis Bay and off northern Namibia.
During the past decade Cape Town-based planktologists have focussed much of their attention on copepod and euphausiid ecology, reinforcing field studies with experiments using animals reared in the laboratory. This has led to a greatly improved understanding of feeding requirements, growth rates and how zooplankton adapt to environmental variability in the southern Benguela. Combined field-laboratory experiments have shown that anchovy are size-selective feeders ("biters") rather than filter feeders and derive most of their energy requirements from large copepods and euphausiids. Conversely sardine (pilchard) feed predominantly by filtering - even filtering relatively large zooplankton such as euphausiids. Only when the density of large food organisms is very low does biting become more important for sardine than filtering (Van der Lingen 1994). Whereas the energy costs of filter feeding for anchovy are high (higher swimming speeds result in high respiration), for sardine the opposite applies i.e. the costs for filter feeding remain much lower than for anchovy even at high swimming speeds, while more energy has to be expended to orientate on prey in a biting mode (Van der Lingen 1995). This food partitioning is illustrated diagrammatically in Fig. 13. The consequence of the different feeding behaviour between anchovy and sardine is that sardine do better when small food particles dominate as in stratified waters, whereas anchovy should do better when the sea is dominated by larger particles, such as during turbulent upwelling conditions and might explain why sardine and anchovy appear to undergo alternative phases of dominance in upwelling regions over periods of 30-60 years (Hutchings and Field 1997). It might also explain some of the observed differences between the southern (rapidly pulsed) and northern (more uniform) parts of the Benguela ecosystem.
The past decade has also witnessed close collaboration between biological oceanographers, physical oceanographers and fishery scientists fostered through joint participation in hydroacoustic fish survey cruises, and this enabled the application of oceanographic knowledge about mechanisms of upwelling and plankton dynamics to be applied directly to solving fishery problems. Apart from obvious increased relevance of research, it also resulted in a greatly improved fundamental understanding of biological oceanographic processes and the role of these as a determinant of fish recruitment. Suites of papers which have appeared in the South African Journal of Marine Science Volumes 5, 12 and 19 and other mainstream international journals document these developments, while progress in various aspects of biological oceanography in the Benguela ecosystem have been very adequately reviewed in a recent article by Hutchings and Field (1997).
Perhaps the most significant finding during recent years is that of Verhehe et al (1998) who observed that crustacean zooplankton abundance, expressed in terms of number of animals, increased by two orders of magnitude in the southern Benguela between 1951 and 1996,suggesting that a major change in the ecosystem has occurred. (This is discussed in Section 5.)
Readers interested in the zooplankton of the Benguela ecosystem within the context of the physics and chemistry and general biology of the South Atlantic Ocean are referred to a recent review article by Boltovskoy
et al (1999).
4.4 Foodweb and carbon budget
As Hutchings and Field (1997) have pointed out, grazing by copepods and euphausiids and the sedimentation of organic material within the Benguela cannot account for the decline in phytoplankton blooms after upwelled water has stabilised, so considerable recycling of organic material must take place in the water column in the southern part of the ecosystem. This has been confirmed by the application of stable isotope techniques (N15 uptake methodology) by Probyn (1992), Waldron
et al (1998) and others that the f-ratio (a relative index of new production) in the Benguela is relatively low (0.2 - 0.3) - only about one half or one third of the value in the Humboldt and California Current systems (Refer also back to Fig. 10). Thus, despite the high rate of new nitrogen input to the shelf waters via upwelling, much of the shelf water is dominated by the microbial foodweb fuelled by recycled nutrients (Hutchings
et al 1995). Following the argument further, as it is new production which determines the productivity at higher tropic levels (fish), and as the food chain in the Benguela has been found to be much more complex (and less efficient) than the classical 20% applicable in a simple short-food chain (phytoplankton® zooplankton® fish), this to some extent explains why the Benguela yields considerably less fish than 100 million tons which simplistic estimates might indicate. Perhaps the most realistic model of the trophic functioning of the Benguela was that developed by Moloney (1992), by incorporating the microbial foodweb. Her model is illustrated diagrammatically in Fig. 14. The fundamental concepts embodied in this simple sketch have major implications for understanding the functioning of upwelling systems and their evident resilience! The understanding of the complexities of the foodweb and underlying processes is an imperative for the management and sustainable utilisation of the living marine resources of the Benguela ecosystem.
In a comprehensive examination of the mechanisms which drive the carbon flux in the Benguela, Monteiro (1996) concluded that upwelling source water moves on to the shelf at three principle sites which are determined by topography (the most important of these being at 27°S near Lüderitz, the others being at 18°S at Cape Frio and at 32°S), and that it is modified on the shelf, becoming enriched in nitrogen and carbon before eventually outcropping at the main upwelling centres (of which there are six). Monterio (1996) hypothesised that the three source sites act as "gates" or barriers to the southward movement of water on the shelf. He developed a model which suggested that the water which upwells at sites north of Lüderitz resulted in outgassing of carbon dioxide, while at Lüderitz and further south, the "carbon pump" and biological activity result in that part of the system being a carbon dioxide sink. Based on certain assumptions about dissolved organic carbon, Monterio calculated that the Benguela system as a whole was a small carbon dioxide sink of 0.34 - 1.5 million tons C/y. The better quantification of upwelling systems such as the Benguela as sources or sinks of carbon dioxide has important implications for the global carbon budget. The Achilles heel remains inadequate knowledge about fluxes of dissolved organic carbon.
5. ENVIRONMENTAL VARIABILITY
A complex array of processes affect the Benguela over a broad spectrum of space and time scales, ranging from the molecular level and fractions of a second to those which span several thousands of kilometers (basin-wide and global) and take place over several months, years, decades or even longer.
There are a number of physical factors which are important determinants of the structure and functioning of the ecosystem, and of these the wind - and in particular the periodicity in the longshore upwelling producing wind - is of over-riding importance. The wind field and wind frequency significantly influence coastal trapped shelf waters (these are internal waves in the sea, referred to earlier), upwelling, formation of fronts, production of filaments, surface and near-surface currents and the depth of the thermocline
inter alia, and a host of dependant chemical and biological processes. The large-scale wind field is also an important determinant of basin-wide circulation, and consequently changes in winds even thousands of kilometers away can impact on the movement of water at the boundaries of the Benguela and within the system.
Apart from winds, seasonal change in insolation (solar heating) is important, influencing the temperature of the surface layer, its thickness and thermocline formation, while variations in light intensity likewise are important in terms of photosynthesis and primary production. What should be recognised is that over one third of the Benguela upwelling area and all of Angola's coastal oceanic waters lie within the tropics, and consequently receive high levels of thermal radiation and light. Tides and tidal currents are important in the near-shore environments, especially in semi-enclosed bays, although in comparison with coastal areas in those parts of the world which experience large tidal ranges, their effects within the Benguela are relatively minor. (The tidal range in the Benguela is typically 1-2m.) There are a number of similarities between the variability in the Benguela and other upwelling systems, but there are also some important differences. What distinguishes the system from other eastern boundary regions is the existence and variations in the northern and southern boundaries viz. the Angola- Benguela front and the Agulhas retroflection area. In addition, the variability in the southern Benguela caused by the free passage of westerly winds and low pressure systems south of the sub-continent make this area quite unique.
However, like in other upwelling systems which are pulsed, the biota in the Benguela ecosystem are generally well adapted to the inherent variability in physical forcing on seasonal and shorter time scales. The biota are, however, less well adapted to sustained major events or changes which occur less frequently i.e. every several years or even over decades. Accordingly, we provide only abridged comments on aspects of small-scale and seasonal variability, and focus most of the discussion on the "catastrophic" occurences which have system-wide impacts. We also highlight some long-term and decadal changes which have been observed in the system.
5.1 Small-scale variability
Processes which occur on time scales of hours to several days and space scales of meters to tens of kilometers are characteristic of upwelling events. These "event scale" processes attracted considerable research attention in the 1970s and 1980s in the southern Benguela, and much of this work has been reviewed by Shannon (1985), Chapman and Shannon (1985) and Shannon and Pillar (1986). The application of remote sensing using aircraft and satellites during wind events in combination with in situ measurements using ships, provided knowledge about the initiation of upwelling, the growth and decay of upwelling plumes off the Cape Peninsula and Cape Columbine following the onset, intensification and subsequent reversal of the upwelling causing winds. Measurements in situ over hours and days provided a good understanding of biochemical processes and plankton dynamics. For example, by following patches of water and frequent sampling of these during upwelling events, the supply and depletion of nutrients, effects of light and light limitation, development of plankton "blooms", species successions, decay processes etc were observed and quantified (e.g. Barlow 1982, Brown and Hutchings 1985). Although most of this research was conducted in the southern Benguela, scientists from the former German Democratic Republic undertook off central Namibia in 1976 one of the most intensive studies of the system over a period of a few weeks, using a combination of repeated surveys along a line of stations and quasi-continuous sampling at a fixed station. (e.g. Postel 1982). Their work showed vividly how the chemistry and biology of the water column changes in response to changes in its physical structure. A somewhat similar experiment was conducted in St Helena Bay a decade later and the findings were published in a special issue of
Progress in Oceanography (Volume 28).
There is so much information contained in the hundreds of scientific publications which resulted from the era of event-type process studies in the southern Benguela, that it is just not possible to condense it adequately and explain it simply. What is apparent, however, is that there exists as superb body of information on the dynamics of upwelling and its consequences in the southern Benguela. Volumes 5 and 12 of the South African Journal of Marine Science published in 1987 and 1992 respectively contain many of the relevant papers. The small and mesoscale processes in the northern Benguela and in Angolan waters have, however, with few exceptions, received less attention. As the systems there function differently from that in the south, simple extrapolation of results and application of some of the concepts developed in the south to the Namibian and Angolan systems may be inappropriate and misleading.
5.2 Seasonal changes and intra-annual variability
The large scale atmospheric weather systems over the south Atlantic display a seasonal pattern of northward and southward migration, and the effect of this seasonal cycle together with changes in insolation, manifests itself in the upper layer. The seasonal changes in longshore upwelling wind stress have already been commented on in Section 2.2 (refer also to Fig. 2). Stratification also plays an important role in seasonal behaviour, for example in the generation of internal tides and in the transport of water. Stratification in the surface layer is intensified by the inflow of fresh warm water from the Congo River, which drains much of central Africa. It seems probable that the Congo River exerts a significant influence on the surface waters off Angola, as the presence of Congo River water can be detected over distances of 1000km or more from its mouth (Dr M. E. L. Buys, pers comm). Information about the seasonal and interannual nature and impact of this water along the coast of Angola may be provided by an examination of the 30 year long record from the monitoring station at Lobito and from other records.
Off Namibia and Angola, there is a distinct seasonal cycle in surface and upper layer temperature, with a maximum in March and a minimum in August/September. The range in seasonal average sea surface temperature is about 4° - 6°C . In the southern Benguela the upwelling season coincides with the period of maximum insolation, and accordingly the range in seasonal average temperature is only 1°-2°C. At the boundaries of the upwelling system, the water column becomes strongly stratified during the austral summer, particularly so over the eastern Agulhas Bank and off Angola, north of the Angola- Benguela front (see Fig 6). The Angola-Benguela front migrates seasonally between its northernmost location near 14°30'S in August and its southernmost position at 17°S in March, the latter being associated with a poleward push of saline Angolan Current water. In the south, the boundary at the Agulhas retroflection likewise displays apparent seasonality, being furthest south in autumn (Dr C. A. Villacastin-Herrero, personal communication). The Agulhas retroflection area is one of the global "hot spots" in terms of variability of temperature and water movements. Apart from the impact of this variability on the South-east Atlantic and the Benguela, it is also important for global climate, as the area south of Africa acts as a veritable "choke point" or "valve" for the movement of heat and salt between the Indo-Pacific and the Atlantic Oceans. (Most of this heat is transported via the six or so rings which are shed each year from the Agulhas Current, so changes in the frequency of ring formation can have major implications for climate)
Although the seasonal changes evident in the upper layer correspond to the seasonal shifts in the weather patterns and insolation, there are no obvious comparable changes in currents. Indeed, the currents near the bottom over the shelf display much less seasonal variability than that which occurs during "events" on 3 to 10 day time scales. There is, however, some suggestion of seasonality in the southward extent of the oxygen depleted shelf water, which reaches a maximum in summer/autumn.
The abundance of phytoplankton throughout the Benguela tends to be higher in summer and autumn, and zones of highest concentration usually lie further offshore during the season of upwelling. Taking the Benguela as a whole, the productive area appears to be larger during summer and autumn than in winter and spring. Subsurface chlorophyll maxima are common throughout the Benguela, particularly marked during summer and autumn when water over the Agulhas Bank is strongly stratified. Storm mixing in areas such as the Agulhas Bank and other shallow parts of the shelf can deepen and erode the thermocline to such an extent that during some winters there is a complete destruction of the thermocline, and the water column becomes well mixed from top to bottom. When this happens higher concentrations of nutrients and consequently higher concentrations of phytoplankton biomass and production can result.
Seasonal trends in zooplankton abundance tend to follow the phytoplankton cycle. Perhaps the most comprehensive record which exists was that provided by Andrews and Hutchings (1980). These authors monitored several parameters in the water column along a line of stations off the Cape Peninsula monthly over three years, and showed that there was about a twofold increase in zooplankton biomass during the upwelling season in the southern Benguela compared with winter (typically 3g dry wt/m2 in January and 1.5g/m2 in August). Further north in the Benguela, there appears to be less seasonality in zooplankton abundance. What is also clear is that consumption of zooplankton by some fish species can significantly reduce zooplankton abundance - even result in "holes" in the distribution on occasions, providing clear evidence that at times food in the form of zooplankton can be a limiting factor for fish.
5.3 Interannual variability and episodic events
There are few reliable and long-term data series from the Benguela region. The longest of these is for sea surface temperature (SST) and it dates from the early part of the 20th century. The majority of the records of other physical, chemical and biological oceanographic parameters are either shorter or are otherwise fragmented in space and in time. A consequence of this is that much of our present understanding of interannual variability in the Benguela has been deduced from case studies of extreme or episodic environmental events, in particular those which significantly impacted on fish abundance and distribution. The most obvious of the extreme events which have occurred in the Benguela are Benguela Nińos (manifest by major incursions of warm tropical water from Angola and from the west into the northern Benguela which occur on an average every ten years), large scale hypoxia in subsurface and bottom water on the continental shelf, anomalous large scale flooding of the southern Benguela by warm Agulhas Current water, major intrusions of cold Sub-Antarctic Water in the south, and sustained anomalous upwelling (or the absence thereof) caused by changes in regional winds. These major environmental "perturbations" will be discussed presently.
A substantial amount of year-to-year environmental variability has been observed in the Benguela ecosystem during the past two decades. The principal changes and fluctuations which have been documented since 1980 can be summarised as follows:
-
Below average sea surface temperature in the shelf area of the northern and southern Benguela during 1982 and 1983
-
A short-lived warm event in the extreme southern Benguela during early 1983, associated with the large
El Nińo which occurred in the Pacific that year, and the commencement then of an extended period of weaker than normal equatorward winds near Cape Columbine (33°S). The period 1982-1983 saw an abrupt change of 30° in the wind direction on the south coast which served to redress long-term changes in the wind direction preceding the event
-
The 1984 Benguela Nińo that followed the cool period in the northern Benguela (but which had little impact on the southern region).This resulted in reduced upwelling off Namibia, the intrusion of warm, saline tropical water from Angola and offshore, deepening of the thermocline and changes in plankton and fish
-
A warm anomaly in the south in 1986, which coincided with the intrusion of Aghulhas Current water into the South-east Atlantic, especially during the winter. This resulted in favourable environmental conditions (short lived) for pelagic fish
-
An onset of cooling in 1987, i.e. a reversal of the warming trend with the influx of Sub-Antarctic water into the southern Benguela in early 1987
-
A period of warming in the southern Benguela in late 1988 and early 1989, evidently associated with changes in the Agulhas retroflection, and a short-lived intrusion of warm water
-
Sustained cooling of shelf waters in the southern Benguela from autumn 1989, and the termination of the negative equatorward wind anomaly (at 33°S) which commenced in 1983. Period of two months of below average reversal of flow at a current meter site at 33°S, completely uncharacteristic of the record, occurred during winter 1989. The abundance off the west coast of the copepod Calanus agulhensis (endemic to the Agulhas Bank) decreased between 1988 and 1991
-
An extended period of below-average trade winds in the extreme south that commenced in the latter part of 1990
-
An eighteen month long cool period commenced in the extreme northern Benguela in mid-1991, with pronounced upwelling off northern Namibia and Angola
-
Intrusion of an Agulhas ring during November and December 1992 adjacent to the coast near Cape Point
-
Anomalously strong south-easterly winds in the southern Benguela during the summer of 1993/1994 which resulted in strong upwelling along the west and south coasts, and coinciding with a widespread negative SST anomaly in the South Atlantic
-
Development and poleward propagation of major hypoxia on the shelf over the full extent of the Benguela region and the occurrence of "black tides" during 1994
-
Benguela Nińo during 1995 in the northern and central Benguela, characterised by poleward flow and widespread warming and altered fish distribution and abundance
-
Return to more normal environmental conditions in the northern Benguela in 1996 and 1997
-
A large intrusion of Agulhas water into the southern Benguela during the summer of 1997/1998, followed by a large bloom of red tide organisms in the St Helena Bay area, localised hypoxia, and an odour of hydrogen sulphide along the coast as far south as Cape Town
The term "Benguela Nińo" was coined by Shannon
et al. (1986) and refers to large scale episodic warm events that occur along the coast of southern Angola and Namibia every ten years on average, and which have a character not unlike the
El Nińo in the Pacific Ocean. Every few years the tropical eastern Atlantic becomes anomalously warm as a consequence of relaxation in the trade winds and the deepening of the thermocline and reduced loss of heat from the ocean to the atmosphere. Occasionally, every ten years on average, this warming is even more extreme, evidently as a consequence of a sudden relaxation of the winds off Brazil, and when this happens the warm water anomaly in the tropical Atlantic travels eastwards and southwards, trapped (guided) by the coast of Africa. The result is a large southward displacement of the Angola-Benguela front, and a flooding of the Namibian shelf by warm tropical water - sometimes very saline (1984), at other times low surface salinity (1995) - depending on the orientation of the flow and the amount of fresh water from the Congo River present. Benguela Nińos are accompanied by increased oxygenation of subsurface and bottom shelf waters either as a consequence of reduced deep flow of water southwards from Angola, or (more likely) reduced primary production and decay on the Namibian shelf. Benguela Nińos are less frequent than Pacific
El Nińos. Just like in the Pacific they are superimposed on the seasonal cycle, but unlike the Pacific the seasonal changes in the Atlantic much larger, and the Atlantic or Benguela Nińos, have a smaller signal. Benguela Nińos occurred in 1934, 1949, 1963, 1984, 1995 and probably around 1910, in the mid-1920s and in 1972-74. The most recent event and its biological impact has been well documented by Gammelsrfd
et al (1998). Benguela Nińos are not necessarily in phase with Pacific El Nińos, but they do appear to be a regional response to changes in the global atmosphere-ocean system.
Just as Benguela Nińos are associated with extreme disturbance of the Angola -Benguela front, extreme disturbances in the retroflection (turning back) of the Agulhas Current at the southern boundary of the Benguela can be manifest as a major incursion of Agulhas water moving into the Benguela system around the Cape of Good Hope. These intrusions may be in the form of shallow surface filaments of warm water (the more usual case), but on occasions warm rings shed from the Agulhas Current can take a more northerly path than usual and impact on subsurface and deeper currents along the edge of the continental shelf - as was the case in 1989. A well documented Agulhas intrusion occurred in 1986 (Shannon
et al 1990). Previous large-scale intrusions may have occurred in 1957 and 1964. The most recent event took place during the post austal summer (1997-1998).
Whereas Agulhas intrusions result in the input of anomalously warm water into the Benguela, incursions of cold Sub-Antarctic water do occur, and occasionally the effect of these can be felt as far north as 33°S (north of Cape Town), as was the case early in 1987. It seems that these rather unusual events are associated with the shedding of rings at the Agulhas retroflection, and they have an appearance of a compensatory northward flow from the Sub-tropical Convergence following the formation of a ring. Their biological consequences in the Benguela are unknown.
Changes in the strength and direction of longshore winds can, if prolonged over several months or longer, result in periods of anomalously low temperatures along the coast (increased upwelling) or warmer conditions associated with more quiescent conditions and downwelling. For example, the abnormally cool shelf waters present in the northern Benguela in the early 1980s were associated with a prolonged period of stronger than normal longshore winds. Cool conditions were again characteristic of the early 1990s off Namibia, while in the southern Benguela in 1993/94 the enhanced upwelling was a consequence of stronger south-easterly winds that summer. Perhaps the most dramatic changes occur on the south coast where surface temperatures lower than 10°C can result on occasions following periods of anomalous easterly (longshore) winds. These conditions tend to occur during times of Pacific La Nińas, while during
El Nińos, westerly winds tend to dominate the Agulhas Bank system and result in downwelling at the coast.
During some years, the most recent being 1993-1994, the oxygen depletion of shelf waters in the Benguela is unusually severe, and this results in widespread anoxia and hypoxia in the system. Although most pronounced in the northern Benguela, episodic depletion of oxygen does occur in the southern Benguela (e.g. in autumn of 1994). These large scale hypoxic and anoxic conditions appear to coincide with quiescent conditions which follow periods of sustained and enhanced upwelling - as a consequence of increased primary production and subsequent decay of phytoplankton blooms (e.g. red tides). It also appears likely that changes in the composition and flow of subsurface waters, in particular the concentrations of dissolved oxygen in/and the southward moving undercurrent along the west coast may be contributing factors, and this suggests that processes taking place off Angola may exert an important influence on the Benguela ecosystem via advection (currents). Irrespective of the causative mechanisms, large scale hypoxia and anoxia result in massive mortalities of marine organisms and changes in distribution and abundance of fish such as hakes (e.g. Hamakuaya
et al 1998). The spectacular impact of such events on non-swimming benthic animals e.g. rock lobsters, is shown in Fig. 16. In the northern Benguela the widespread occurrence of oxygen depleted/deficient water does not coincide with Benguela Nińo events, but appear to precede these.
5.4 Decadal changes and regime shifts
There is a growing body of evidence which suggests that marine ecosystems undergo decadal-scale fluctuations driven by variability in climate. This is most apparent at the higher trophic levels (fish). For example in the Humboldt Current system decadal period switches in dominance between sardine and anchovetta have occurred this century, and it is also apparent from the sediment record (from analysis of fish scale deposits) that species alternations have occurred during past centuries i.e. prior to the advent of fishing and other anthropogenic impacts. Moreover it has been demonstrated that populations of sardine in different parts of the Pacific (off Chile/Peru, California/Mexico and Japan) have undergone synchronous fluctuations this century. In the Benguela ecosystem there have been corresponding fluctuations in sardine abundance, although these appear to be out of phase with those in the Pacific. Also, there is evidence in the southern Benguela, at least, of species alternations-regime shifts- between anchovy and sardine, and in the northern Benguela the sediment record suggests that both species undergo decadal-scale fluctuations, at times alternating species dominating, at others the two species fluctuating synchronously (e.g. Shackleton 1986). What is also apparent from the sediment record is that there can be long periods (several decades) where both species are absent or only present at a low biomass.
As previously indicated, there are few long-term data series of environmental parameters for the Benguela region. The available indices do, nevertheless, show that changes and fluctuations on decadal or longer time scales have occurred in the Benguela this century, and superimposed on this variability a progressive warming of surface waters of about 0.7°C from 1920 is apparent throughout the Benguela and South-east Atlantic (This value has been corrected for changes in measuring instruments). The analysis of Taunton-Clark and Shannon (1988) showed that the 1920s and 1930s were cool years in the region and that a change to warmer conditions took place during the 1940s, followed by a gradual strengthening of the south-easterly trade winds over the next few decades. A system-wide change occurred in the late 1960s, to be followed by an extended warm period. The warming trend accelerated during the 1980s and this decade was the warmest this century in the South-east Atlantic. Longshore wind stress in the Benguela increased sharply after 1974 and the shelf waters along the west coast of South Africa and Namibia were abnormally cold in the early 1980s. In the southern Benguela (at least), wind record displays a sharp change in 1982/1983, with significantly lighter winds blowing for the remainder of the decade (Shannon
et al 1992). It is perhaps significant that there is substantial decadal-scale variability evident in the post 1960 winds off Brazil (Carton and Huang 1994). The Gulf of Guinea temperature records also suggest an apparent periodicity between large sustained warm events of about 10 years.
While the physical mechanisms linking decadal variability observed in the South-east Atlantic with that of the Indo-Pacific and North Atlantic is not understood, it is likely that the Benguela is influenced by external changes, and must be viewed within the context of the global ocean-atmosphere system. Generally, warming/cooling in the tropical Atlantic appears to be out of phase with that in the Pacific. What is also quite conceivable is that changes in the formation of Deep Water in the North Atlantic - such as occurred in the late 1960s during the "great salinity anomaly" will alter the northward movement of heat through the Atlantic and the flux of water and heat south of Africa. (Note that the South Atlantic is the only ocean where the net movement of heat is towards the equator, and much of this comes from the Indo-Pacific.)
While decadal-scale changes are evident from some of the physical parameters and in fish populations, these are not so apparent in the chemistry and plankton because of the fragmented nature of the available records. The available information does however point to the likelihood that comparable changes in the plankton has taken place, and by extrapolation from the biology and physics, that changes in nutrient supply have also occurred. Brown and Cochrane (1991) analysed chlorophyll a (a proxy for phytoplankton biomass) data available for the southern Benguela between 1971 and 1989. Although there appears to be a decreasing trend during the two decades from about 3.5 mg/m3 to 2.0 mg/m3, it is not statistically significant because the data are highly patchy. In what is perhaps the most important paper on zooplankton in the Benguela during recent years, Verheye
et al. (1998) found that the abundance of animals in all main toxonomic groups in the St Helena Bay area increased by at least ten fold between 1951 and 1996 (These measurements applied to the main pelagic fish recruitment season viz March-June). Total zooplankton abundance expressed in numbers of animals increased by more than one hundred fold. The increase was accompanied by a significant shift in the community structure of near shore zooplankton, with a trend towards smaller size organisms. Whether the observed change is a consequence of reduced predation pressure by anchovy and sardine ("top down control") or climatologically induced viz - upwelling, primary production and entrainment ("bottom up control"), is not clear at this stage. Other changes in zooplankton include the decrease in abundance on the west coast of the copepod Calanus agulhensis between 1988 and 1991 (previously mentioned) and the relative scarsity of the euphausiid Nyctiphanes capensis now in comparison with the 1950s (E. lucens is currently dominant).
5.5 Recent developments
In March/April 1998 environmental variability in the South-east Atlantic was the focus of an important International Symposium and Workshop, held in Swakopmund, Namibia. Hosted by the Namibian Ministry of Fisheries and Marine Resources and sponsored by The World Bank, BENEFIT, the German Agency for Technical Cooperation (GTZ), the Royal Norwegian Consulate General in Windhoek (Office for Development Cooperation) and the Scientific Committee for Oceanic Research (SCOR) of the International Council for Science (ICSU) provided a showcase of current knowledge and ideas about environmental variability in the Benguela, possible linkages/teleconnections with variability elsewhere in the Atlantic Ocean and with the global system, the time and space scales of the variability and ecosystem implications. Ninety leading scientists participated in the Symposium/Workshop. Of these 30 were from overseas and another 5 were international experts seconded to-or resident in- the SADC region, making this meeting among the most international of any oceanographic conference held in southern Africa. Comprehensive proceedings are available (Shannon and O'Toole 1998).
6. ISSUES, PROBLEMS, THREATS AND GAPS IN KNOWLEDGE
The different, but complementary, processes associated with the development of BENEFIT and the BCLME, as well as national strategic planning initiatives have generated a wealth of information about the main environmental issues, problems and threats in the Benguela region and gaps in knowledge and understanding. Various reports e. g. Anon (1997), Hempel and Haslund (1997), Shannon and O'Toole (1998) and Croll (1998) document these fairly comprehensively. Accordingly, the following sub-sections draw on the available literature, although obviously we have had to synthesise and provide a degree of re-interpretation of the various views and perceptions.
6.1 Fundamental issues
As pointed out earlier in this Overview, the Benguela differs from other upwelling ecosystems in several respects, e. g. it is bounded on northern and southern ends by warm- water regimes and strong fronts are associated with these boundaries; in the southern Benguela upwelling is strongly pulsed on a time scale of 3 - 10 days due to the free passage of low pressure systems moving past the African continent; the Agulhas retroflection results in a high degree of complexity and variability which does not have an equivalent in any other upwelling area, and results in a net movement of heat in the South-east Atlantic towards the equator and conduit linking the Benguela with the Indo-Pacific; the continental shelf is very variable in depth and width; a key fish spawning area exists upstream of two important upwelling centres and planktonic fish and larvae utilise a strong jet current to move to the recruitment area; the northern, central and southern parts of the Benguela function very differently, with the peak upwelling seasons in the north and south being distinctly out of phase, either in synchrony with , or opposing, the annual solar-heating cycle. The complex and unique character of the Benguela thus necessitates special attention. Ideas and principals developed elsewhere will not necessarily be simply transferable and applicable to the Benguela.
To date little attention has been given to transboundary environmental issues and the management of these. This is largely due to the nature and location of the external and internal system boundaries. The northern boundary of the main upwelling system viz the Angola -Benguela front lies close to the country boundary between Angola and Namibia, while the major "internal boundary" feature viz the Lüderitz upwelling cell which effectively divides the Benguela into northern and southern parts is not far from the Namibia-South Africa border. The colonial past, coupled with the distinct distributions of the principal harvested living marine resources has resulted in the countries in the region focussing activities within their own EEZs. Moreover the civil war in Angola and political problems in the region during the 1970s and 1980s resulted in scant attention being given to cross-system boundary environmental issues. A question which needs to be addressed within the context of the BCLME is definition of the northern boundary of the Benguela Current large marine ecosystem per se. Perhaps the term Greater Benguela Current System might be appropriate as this would clearly include the Angolan region and the Agulhas retroflection?
Management of Benguela living and non-living resources has in the past not been integrated adequately either within countries or within the region. There has also been a general lack of an ecosystem approach to the management of living marine resources, although attempts have been made to take environmental considerations into account, for example, in setting catch levels in the different countries. The real challenge for the BCLME initiative is the development of viable integrative environmental mechanisms for the region as a whole i.e. at the ecosystem level.
6.2 Environmental variability
Although the Benguela environment displays a high degree of variability across a wide range of time and space scales, there is a growing realisation and consensus that it is the major sustained events and changes which seem to impact on the ecosystem as a whole. From a management point of view, there is a need to predict these large environmental events and changes, and the impacts of these on the system. There is evidence that the Benguela responds to basin-scale processes, in particular changes in the physical environment in the tropical Atlantic (e.g. changes in windstress off Brazil) and in the North Atlantic (e.g. North Atlantic Oscillation), changes in the Southern Ocean (e.g. Antarctic Circumpolar "Wave") and to those in the Indian Ocean (variations in winds, the flow and leakage of the Agulhas Current). These changes are in turn are linked to global ocean-atmosphere processes. In other words, the Benguela must be viewed within the context of the global environment, not in regional isolation. However, the problem is that the teleconnections and linkage processes are not properly understood. Nevertheless, documented changes in the state of the ecosystem and regime shifts are congruent with decadal scale changes which have taken place in other ecosystems, but again it is difficult to distinguish between a natural environmentally driven cause (bottom up) and that caused by humans (top down). At the First Regional BCLME Workshop, the changing environment was a key issue raised within all three working groups. What is very clear, however, is that predicting environmental variability and change and the impacts is likely to be extremely difficult - yet so important. It will necessitate comprehensive monitoring, process studies and modelling at the very least.
While it is apparent that major environmental events such as system-wide hypoxia/anoxia and Benguela Nińos as well as changes in primary and secondary production, occurrences of harmful algal blooms (red tides), changes in currents etc are important determinants or indicators of ecosystem variability and change, the present level of environmental monitoring within the Benguela region is totally inadequate to document these -
let alone to use the information for any predicting or forecasting. The issue of environmental monitoring in the Benguela was addressed in some detail at an International Workshop held in Swakopmund in April 1998, and a monitoring framework was developed (mainly for Namibia). Implementation of a cost-effective regional monitoring strategy and system along these lines is seen as an important component of the BCLME programme. However, environmental monitoring will be expensive and is not readily automated. Although obviously satellite remote sensing will comprise an essential element of the environmental monitoring, it will require appropriate in situ measurements ("ground truthing" or validating) to be useful.
Some of the more obvious features/issues re environmental variability in the Benguela e.g. harmful algal blooms and the role of sulphur in the ecosystem (cf sulphur "eruptions") will be addressed at specialist international workshops which are scheduled to take place in Namibia within the next six months. The possible expansion of a monitoring buoy network from the tropical Atlantic to the Benguela (PIRATA array) is being examined.
The introduction into the ecosystem of exotic species of marine organisms accidentally e. g. via ballast water discharges from ships entering ports, or deliberately e.g. unauthorised mariculture ventures, is a potential threat, but remains unquantified. Perhaps the biggest threat to the Benguela is from the cumulative effect of small changes, some of which may be imperceptible, as a consequence of human activity in the region (fishing, mining etc) and as a consequence of global climate change.
6.3 The Benguela and global environmental (climate) change.
There are two issues here, viz the role of the greater Benguela ecosystem in global climate change processes, and the impact of climate change on the Benguela. With respect to the first issue, there is some uncertainty as to whether the Benguela is a net source or sink of atmospheric carbon dioxide (see Section 4) with the "physical pump" supplying CO2 through outgassing from upwelling opposing the "biological pump" which will remove CO2 from the atmosphere through primary production and the marine food web. Work of Monteiro (1996) indicates that the southern Benguela may be a net sink and the northern Benguela a net source, with the Benguela as a whole being a small sink. However, if the Angola region and Angola Dome were to be included, the picture may alter radically. Similarly changes in primary production and the marine food web could further alter the balance in future. Thus knowledge of what happens in the Benguela and other upwelling systems is important for global models predicting future climate change. A problem is that very few studies have been made in the Benguela on the flux of carbon, and this is exacerbated by the dearth of marine chemists in southern Africa and the low priority given by regional funding agencies to marine chemistry.
The importance of ocean processes in climate studies and prediction is highlighted by the fact that the top 1 metre of the sea contains as much heat as the entire 50km overlying column of atmosphere! More-over, the South Atlantic is the only ocean in which there is a net movement of heat from the south to the equator. This is in part due to the "leakage" of warm Agulhas Current water into the South Atlantic via the Agulhas retroflection "choke point" or "valve". This area is part of a global ocean heat "conveyer belt" linking the Pacific, Indian and Atlantic Oceans which has a return cold conveyer arm deeper down. This is illustrated schematically in Fig 17. When global climate warms the conveyer speeds up, when it cools it slows down. Thus the marine environmental processes which take place in the southern part of the greater Benguela system viz in the Agulhas retroflection may area have an enormous impact on world climate. This is recognised internationally, and highlights the global importance of the Benguela environment - the conduit for the heat, a veritable global climate valve.
Global climate has warmed by about 0.8°C this century, and the balance of evidence suggests a discernable human influence on global climate. If it is accepted that significant change will occur in global climate in the next century, this is likely to impact on the Benguela - not so much through small increases in ocean temperature and sea level (expected to increase by about 50cm by the end of the 21st century) but through changes in wind over the South Atlantic and Indian Oceans. Coastal winds are likely to strengthen and if this happens, upwelling will intensify and the present upwelling area may expand. This in turn would have consequences for primary production and the food web and also for global climate. In addition there may well be impacts from increased levels of ultraviolet radiation resulting from the destruction of stratospheric ozone by CFCs. (In this respect the "ozone hole" is still expanding).
There remains so much uncertainty about the likely changes and interactions, that any scenarios developed at this stage would be purely speculative. Nevertheless it would be reasonable to anticipate that ecosystem changes and regime shifts will occur during the 21st century and that these changes are likely to impact on the economies of Angola, Namibia and South Africa. This again highlights the need for appropriate environmental monitoring, studies of cause and effect and modelling with the view to improving predictability. However, the impact of changes in the Benguela marine environment may be dwarfed by the likely impact of climate change on rainfall in southern Africa and of this on the economy of SADC countries.
6.4 Gaps in knowledge and understanding
Some of the more obvious shortcomings or gaps are listed below in point form. (These are not in any particular order)
-
Teleconnections between the Benguela and the tropical Atlantic, North Atlantic, Southern Ocean and Indo-Pacific and the linkage mechanisms (cf global weather patterns)
-
Predictability of environmental change and ecosystem response
-
Dynamics of formation, advection and impact of oxygen deficient/depleted water in the greater Benguela system (hypoxia/anoxia phenomena)
-
Dynamics of the Angola Current, its variability, and associated processes
-
Role of the Walvis Ridge - physical, chemical and biological
-
Various atmosphere-water-sediment chemical processes
-
Angola - Benguela front dynamics and stability and biological impact thereof
-
The Benguela undercurrent and spatial continuity
-
Changes in system wide plankton abundance and species composition, during the past 50 years. (Samples exist, but require analysis and interpretation)
-
The Benguela sulphur cycle/system and its biological consequences
-
Plankton production in Angola's EEZ and in the Angola Dome, and associated biochemical processes
-
Harmful algal blooms (red tides) - distribution, frequency, species,impacts
-
Extent and impact of exotic species of marine flora and fauna introduced from ballast water discharges (control measures?)
-
Seasonal and interannual variability of the Agulhas Current and Agulhas retroflection
-
Extent of marine pollution in the Benguela and its impact
-
Impact of climate change on the Benguela ecosystem
-
Quantification of the greater Benguela as a net source or sink of carbon dioxide
-
Environmental (ecosystem) impact of offshore mining activities and oil/gas exploration/extraction
-
Transboundary environmental policy/management
-
Human-environment-resource interactions : quantification of natural environmental variability vs human impact on the ecosystem
6.5 Infrastructure and human capacity
Comprehensive information about human capacity and infrastructure available in Angola, Namibia and South Africa and shortcomings is provided in Appendix I. The principal constraints and problems are briefly as follows:
-
There is a steep gradient from south to north in terms of both expertise and infrastructure. In the case of Angola, while laboratory and office accommodation is good, there is a general lack of laboratory equipment and instruments and the necessary support infrastructure. There are few scientists dedicated to environmental studies in Angola, with a lack of chemical and biological oceanographers. Namibia has excellent facilities but few posts are allocated to environmental research and monitoring.
-
Except in South Africa, there is a general lack of trained and experienced technicians to operate and support (service/maintain) high technology equipment and to provide the necessary technical support to research staff. This shortcoming has serious implications for environmental monitoring programmes, and results in inefficient use of oceanographers with interpretive skills
-
There are relatively few scientists in the region with broad-based interpretive skills i.e. who can integrate and synthesise across disciplines. There is also a shortage of modellers, although this is being addressed by South Africa
-
There are very few marine chemists and chemical technicians in the region - including in South Africa. Marine chemistry is de facto the "Cinderella" activity in the Benguela
-
While there is a need throughout the region to develop oceanographic expertise, in particular in Angola but to a lesser extent also in Namibia and South Africa, there is a shortage of tenured posts available for new graduates from universities and technikons i.e. there are needs, but few available jobs
-
Angola does not have a viable research ship, and relies on assistance from Norway. The situation in Namibia and South Africa is more satisfactory, although funding and manpower constraints in the last mentioned country are making it difficult to operate research ships such as F.R.S Africana (which is now 17 years old)
6.6 Funding
Funding for marine science and technology throughout the region is limited, and is decreasing every year in real terms, because of other more pressing national priorities. In Angola salaries of qualified scientists at IIP are very poor and staff are obliged to seek alternative means to supplement incomes. Environmental research and monitoring is seen in all countries as a lower priority than e.g. resource based work as it has a longer "pay back" period (cf pure research vs applied research). As a consequence funding for marine environmental research and monitoring in the Benguela is inadequate - even in South Africa. Unless the funding problem can be addressed adequately, and also flexibility to utilise funds improved, there will be little prospect of a viable environmental component within the
BCLME.
7. ACKNOWLEDGEMENTS
The authors are most appreciative of the help and encouragement provided by the various experts consulted during the preparation of this overview. In particular we wish to acknowledge with sincere thanks support from staff of the governmental lead agencies in the three countries participating in the Benguela Current Large Marine Ecosystem Programme viz. Ministry of Fisheries and Marine Resources (Namibia), Instituto de Investigaçao Pesqueira (Angola) and Sea Fisheries Research Institute of the Department of Environmental Affairs and Tourism (South Africa).
This Overview was funded by The World Bank via a Block B grant from The Global Environmental Facility (GEF) with The United Nations Development Programme (UNDP) as implementing agency. Assistance provided by UNDP staff, both in New York and Windhoek, is acknowledged with appreciation. The work was guided by the BCLME Management Committee, the Members of which provided useful comments and helpful advice.
The Benguela-Environment-Fisheries-Interaction-Training Programme (BENEFIT), a regional initiative of Angola, Namibia and South Africa which was launched in 1997 was a catalyst for the development of the BCLME Programme. This Overview draws on information and ideas generated by the embryonic BENEFIT, and from the regional and international oceanographic experts who have participated in the initiation of BENEFIT.
Valuable information on the Angolan component of the Benguela environment was provided by Messrs. V. L. L. Filipe and F. Pereira, of the IIP in Luanda, while a recent report prepared by Drs G. Hempel and O. H. Haslund on the status of marine science in Angola was an important source of information about Angolan institutional capacity. Dr H. Marques, Advisor at IIP, Luanda, provided many useful comments and suggestions on the draft text which has been revised accordingly.
Satellite images of the Benguela were kindly provided courtesy of Ms S. Weeks (OceanSpace CC) and her assistance in this respect is gratefully acknowledged. The majority of the diagrams were generated by Mr A. P. van Dalsen (SFRI) whose considerable cartographic skills are apparent in the illustrations.
8. REFERENCES
During the past decade several thousand scientific publications on the Benguela ecosystem and its harvested resources have appeared in the primary scientific literature, the majority of which dealt with the southern Benguela. A full listing of these is beyond the scope of this document. The key references have, however, been cited in the text and details of these follow. In addition readers are referred to suites of papers in volumes 5,12 and 19 of the South African Journal of Marine Science, (1987, 1992 and 1998 respectively) and in Progress in Oceanography, Vol 28 (1991).
ANDREWS, W. R. H. and L. HUTCHINGS 1980 - Upwelling in the southern Benguela Current. Prog.Oceanogr. 9:81 pp
ANON 1997 - BENEFIT Science Plan, BENEFIT Secretariat, Windhoek, Namibia, 90 pp.
BAILEY, G. W. 1979 - Physical and chemical aspects of the Benguela Current in the Lüderitz region. MSc thesis Univ. Cape Town:225 pp
BAILEY, G. W. and J. ROGERS 1997 - Chemical oceanography and marine geoscience off southern Africa: Past discoveries in the post-Gilchrist era, and future prospects. Trans. Roy. Soc. S. Afr. 52(1):51-79.
BAKUN, A. and C. S. NELSON 1991 - The seasonal cycle of wind-stress curl in subtropical eastern boundary current regions. J. Phys. Oceanogr. 21:1815-1834.
BANG, N. D. and W. R. H. ANDREWS 1974 - Direct current measurements of a shelf-edge frontal jet in the southern Benguela system. J. mar. Res. 32(3):405-417.
BARLOW, R. G. 1982 - Phytoplankton ecology in the southern Benguela Current:3 Dynamics of a bloom. J. exp. Mar. Ecol. 63:239-248.
BOLTOVSKOY, D., GIBBONS, M. J., HUTCHINS, L. and D. BINET 1999 - General biological features of the South Atlantic. In South Atlantic Zooplankton (D. Boltovskoy, Ed) Backhuys Publishers, Leiden, pp 1-42.
BOYD, A. J. 1987 - The oceanography of the Namibian shelf. Ph.D. thesis University of Cape Town:190pp.
BOYD, A. J., HEWITSON J. D., KRUGER, I. and F. LE CLUS 1985 - Temperature and salinity trends off Namibia from August 1982 to August 1984, and their relation to plankton abundance and reproductive success of pelagic fish. Colln scient. Pap. int. Commn SE Atl. Fish. 14(1):85-99.
BOYD, A. J. and F. P. SHILLINGTON 1994 - Physical forcing and circulation patterns on the Agulhas Bank. S. Afr. J. Sci. 90: 114-122.
BROECKER, W. S. 1991 - The great ocean conveyer. Oceanography 4: 79-89.
BROWN, P. C. and L. HUTCHINGS 1987: - The Development and decline of phytoplankton blooms in the southern Benguela upwelling system. 1. Drogue movements, hydrography and bloom development. In Payne A. I. L., Gulland, J. A. and Brink, K. H. (Eds). The Benguela and Comparable Ecosystems. S. Afr. J. mar. Sci. 5:357-391.
BROWN, P.C. PAINTING, S. J. and K. L. COCHRANE 1991 - Estimates of phytoplankton and bacterial biomass and production in the northern and southern Benguela ecosystems. S. Afr. Mar. Sci. 11:537-564.
BUBNOV, V. A. 1972 - Structure and characteristics of the oxygen minimum layer in the Southeastern Atlantic. Oceanology 12(2):193-201.
CALVERT, S. E. and N. B. PRICE 1971 - Upwelling and nutrient regeneration in the Benguela Current, October, 1968. Deep-Sea Res. 18: 505-523.
CARTON, J. A. and B. HUANG 1994 - Warm events in the tropical Atlantic. J. Phys. Oceanogr. 24:888-903.
CHAPMAN, P. and L. V. SHANNON, 1985 - The Benguela ecosystem. 2. Chemistry and related processes. In Oceanography and Marine Biology. An Annual Review 23. Barnes, M. (Ed.). Aberdeen; University Press:183-251.
CHESTER, R. and J. H. STONER 1975 - Trace elements in total particulate material from surface sea water. Nature 255 : 50-51.
COPENHAGEN, W. J. 1934 - Occurrence of sulphide in certain areas of the sea bottom on the South African coast. Investl. Rep. Fish. Mar. Biol. Surv. Div. Un. S. Afr. 3 : 11 pp.
CROLL, P. 1998 - Benguela Current Large marine Ecosystem (BCLME) First Regional Workshop UNDP, 22-24 July 1998, Cape Town, South Africa (Moderator's report on Workshop) : 73 pp.
DE DECKER, A. H. B. 1970 - Notes on an oxygen-depleted subsurface current off the west coast of South Africa. Investl. Rep. Div. Sea Fish. S. Sfr. 84:24pp.
DE VILLIERS, S. 1998 - Seasonal and interannual variability in phytoplankton biomass on the southern African continental shelf : evidence from satellite-derived pigment concentrations. S. Afr. J. mar. Sci. 19 (in press).
DIAS, C. A. 1983 - Preliminary report on the physical oceanography off southern Angola, March and July 1971. Colln scient. Pap. Int. Commn S. E. Atl. Fish. ICSEAF 1983 (Part 11) : 103-112.
EAGLE, G. A., BARTLETT, P. D. and M. V. LONG 1982. Council for Scientific and Indistrial Research, S. Afr. Report 571:79pp
GAMMELSROD, T. 1998 - Global thermohaline circulation and the Benguela Current, climatic implications and impact on biota. In Proceedings of the International Symposium on Environmental Variability in the South-east Atlantic, Swakopmund, Namibia, March-April 1998:5pp.
GAMMELSROD, T., O'TOOLE M. J., BARTHOUSE, C. J., BOYER, D. and V. L. FILIPE 1998 - Intrusion of warm surface water layers at the Southwest African coast in February-March 1995:The Benguela Nińo '95: S. Afr. J. mar. Sci. 19 (in press).
GIBBONS, M. J. 1999 - An introduction to the zooplankton of the Benguela Current region. ISBN 0-620-244225-6 (Published by Ministry of Fisheries and Marine Resources, Windhoek, Namibia) 51 pp.
GORDON, A. L. and K. T. BOSLEY 1991 - Cyclonic gyre in the tropical South Atlantic. Deep- Sea Res. 38 (Suppl. 1A) S323-S343.
GORDON, A. L., BOSLEY, K. T. and F. AIKMAN 1995 - Tropical Atlantic water within the Benguela upwelling system at 27°S. Deep- Sea Res. 42:1-12
HAMUKUAYA, H., O'TOOLE, M. J., and P. M. J. WOODHEAD 1998 - Observations of severe hypoxia and offshore displacement of Cape hake over the Namibian shelf in 1994. S Afr. J. mar. Sci. 19 (in Press).
HART, T. J. and R. I. CURRIE 1960 - The Benguela Current. Discovery Rep. 31: 123-297.
HEMPEL,G. and O. H. HASLUND 1997 - Notes on the State of Marine Science in Angola : Report on a Visit to Angola, 12-17 April 1997. IOC/UNESCO : 17 pp.
HORSTMAN, D. A. 1981 - Reported red-water outbreaks and their effects on fauna of the west and south coasts of South Africa. Fish. Bull. S. Afr. 15 : 71-88.
HUTCHINGS, L., and J. G. FIELD 1997 - Biological oceanography in South Africa, 1896-1996:Observations, mechanisms, monitoring and modelling. Trans. Roy. Soc. S. Afr. 52(1): 81-120.
HUTCHINGS, L., PITCHER, G. C., PROBYN, T. A. and G. W. BAILEY 1995 - The chemical and biological consequences of coastal upwelling. In Upwelling in the Ocean:Modern Processes and Ancient Records. Summerhayes, C. P., Emeis, K-C., Angel, M. V., Smith, R. L. and B. Zeitzschel (Eds). John Wiley & Sons Ltd: 65-81.
KOSTIANOY, A. G. 1996 - Use of satellite data for the analysis of the fronts in the S. E. Atlantic (with as an example the Angola-Benguela Frontal Zone) Issled. Zemli iz kosmosa 4:77-86 (in Russian).
LASS, H.U. and M. SCHMIT 1998 - The Angola-Benguela front - measurements, modelling and consequences. In Proceedings of the International Symposium on Environmental Variability in the South-east Atlantic, Swakopmund, Namibia, March/April 1998 : 13pp. Plus 20pp diagrams.
LUTJEHARMS, J. R. E., MEYER, A. A., ANSORGE, J. J., EAGLE, G. A. and M. J. ORREN 1996 - The nutrient characteristics of the Agulhas Bank. S. Afr. J. mar. Sci. 17: 253-274.
MOLONEY, C. L. 1992 - Simulation studies of trophic flows and nutrient cycles in Benguela upwelling foodwebs. In Benguela Trophic Functioning. Payne, A. I. L., Brink, K. H., Mann K. H. and R. Hilborn (Eds). S. Afr. J. mar. Sci. 12: 457-476.
MONTEIRO, P. M. S. 1996 - The oceanography, the biogeochemistry and the fluxes of carbon dioxide in the Benguela upwelling system. Ph.D thesis, Univ. Cape Town, S. Afr.: 354 pp.
MOROSHKIN, K. V., BUBNOV, V. A. and R. P. BULATOV 1970 - Water circulation in the eastern South Atlantic Ocean. Oceanology 10(1): 27-34.
NELSON, G. 1980 - Poleward motion in the Benguela area. In Neshyba, S. J., Moers, C. N. K., Smith, R. L. and Barber, R. T. (Eds) Poleward Flows along Eastern Ocean Boundaries. Springer, New York: 110-130 (Coastal and Estuarine Studies 34)
NELSON, G. and L. HUTCHINGS 1983 - The Benguela upwelling area. Prog. Oceanogr. 12(3): 333-356.
ORREN, M. J. 1969 - The abundance and distribution of some elements in the ocean Ph.D thesis, Univ. Cape Town, S. Afr. 135 pp plus 50 figures.
ORREN, M. J. 1971 - In The Ocean World, M. Uda (Ed) Proc. Joint Oceanogr. Assembly, Tokyo, 1970). Japan Soc. For Promotion of Science, Tokyo, 187-189.
PAREDES, F. J. 1962 - On occurrence of red water on the coast of Angola. Mem. Jta. Invest. Ultramar. 32(2) : 87-114.
PARRISH, R. H., BAKUN, A., HUSBY, D. M. and C. S. NELSON 1983 - Comparative climatology of selected environmental processes in relation to eastern boundary current pelagic fish reproduction. In Proceedings of the Expert Consultation to Examine Changes in Abundance and Species Composition of Neritic Fish Resources, San Jose, Costa Rica, April 1983. Sharp. G. D. and J. Csirke (Eds). F. A. O. Fish. Rep. 291(3): 731-777.
PEREIRA, A. de F. and V. FILIPE 1998 - Summary of some recent oceanographic and environmental variability information from Angolan waters. Unpublished BCLME Rept. 28pp
PICAUT, J. 1985 - Propagation and seasonal upwelling in the eastern equatorial Atlantic. In Recent Progress in Equatorial Oceanography : A Report of the Final Meeting of SCOR Working Group 47, Venice, April 1981. Mc Creary, J. P., Moore, D. W., White, J. M. (Eds). Fort Lauderdale. Fl., Nova Univ. NYIT Press: 271-281.
PITCHER, G. 1990 - Phytoplankton seed populations off the Cape Peninsula upwelling plume, with particular reference to resting spores of Chaetoceros (Bacillariophyceae) and their role in seeding upwelling waters. Estuarine Coastal Shelf Science 31 : 283-301.
PITCHER, G., AGENBAG, J., CALDER, D., HORSTMAN, D., JURY. M. and J. TAUNTON-CLARK, 1995 - Red tides in relation to the meteorology of the southern Benguela upwelling system. In Harmful Marine Algal Blooms. Lassus, P., Arzul, G., Erard, E., Gentien, P. and C. Marcaillou (Eds). Technique et Documentation - Lavoisier, Intercept Ltd, 1995.
PITCHER, G. and A. J. BOYD 1996 - Cross-shelf and alongshore dinoflagellate distributions and the mechanisms of red tide formation within the southern Benguela upwelling system. In Harmful and Toxic Algal Blooms, Yaumoto, T., Oshima, Y. and Y. Fukuyo (Eds) Paris, IOC/UNESCO : 243-246.
PITCHER, G. C. 1998 - Harmful algal blooms of the Benguela Current. Report published by Ministry of Fisheries and Marine Resources, Windhoek, Namibia. 20pp.
POOLE, R. and M. TOMCZAK 1999 - Optimum multiparameter analysis of the water mass structure in the Atlantic Ocean thermocline. Deep-Sea Res I 46:1895-1921.
POSTEL, L. 1982 - Mesoscale investigations on space-temporal variability of the zooplankton biomass in the upwelling regions of Northwest and Southwest Africa. Rapp. P. -v Reun. Cons int. Explor. Mer. 180:274-279.
PRICE, N. B. and S. E. CALVERT 1977 - The contrasting geochemical behaviours of iodine and bromine in recent sediments from the Namibian shelf. Geochim. Cosmochim. Acta. 41:1769-1775.
PROBYN, T. A. 1992 - The inorganic nutrition of phytoplankton in the southern Benguela: new production, phytoplankton size and implications for pelagic foodwebs. In Benguela trophic Functioning. Payne, A. I. L. , Brink, K. H., Mann, K. H., and R. Hilborn (Eds). S. Afr. J. mar. Sci. 12: 411-420.
PROBYN, T. A., WALDRON, H. N. and A. G. JAMES 1990 - Size-fractionated measurements of nitrogen uptake in aged upwelled waters : implications for pelagic food webs. Limnol. Oceanogr. 35: 202-210.
RENNEL, J. 1778 - A Chart of the Bank L'Agulhas and Southern Coast of Africa.
SHACKLETON, L. Y. 1986 - An assessment of the reliability of fossil pilchard and anchovy scales as fish population indicators off Namibia. MSc. Thesis, Univ. Cape Town, S. Afr. 141 pp.
SHANNON, L. V. 1985 - The Benguela ecosystem. 1. Evolution of the Benguela, physical features and processes. In Oceanography and Marine Biology. An Annual Review 23. Barnes, M. (Ed). Aberdeen; University Press: 105-182.
SHANNON, L. V. (Ed). 1985 - South African Ocean Colour and Upwelling Experiment. Cape Town; Sea Fisheries Research Institute: 270 pp.
SHANNON, L. V., AGENBAG, J. J., and M. E. L. BUYS 1987 - Large and mesoscale features of the Angola-Benguela front. In Payne, A. I. L., Gulland, J. A. and K. H. Brink (Eds) The Benguela and Comparable Ecosystems, S. Afr. J. mar Sci. 5 : 11-34.
SHANNON, L. V. 1989 - The physical environment. In Oceans of Life off Southern Africa, Payne, A. I. L. and Crawford, R. J. M (Eds) Illustrated by van Dalsen, A. P. Vlaeberg, Cape Town, pp12-27
.
SHANNON, L. V., AGENBAG, J. J., WALKER, N. D. and J. R. E. LUTJEHARMS 1990 - A major perturbation in the Agulhas retroflection area in 1986. Deep Sea Res. 37 (3): 493-512.
SHANNON, L. V. BOYD, A. J., BRUNDRIT, G. B. and J. TAUNTON-CLARK 1986 - On the existence of an
El Nińo-type phenomenon in the Benguela system. J. mar. Res. 44(3): 495-520.
SHANNON L. V. and thirteen co-authors 1992 - The 1980s - A decade of change in the Benguela ecosystem. In Benguela Trophic Functioning, Payne, A. I. L., Brink, K. H., Mann, K. H. and R. Hilborn (Eds). S. Afr. J. mar. Sci. 12 : 271-296.
SHANNON, L. V. and G. NELSON, 1996 - The Benguela : Large scale features and processes and system variability. In The South Atlantic Past and Present Circulation. Wefer, G., Berger, W. H., Siedler, G. and D. J. Webb (Eds). Springer Verlag, Berlin, Heidelberg: 163-210.
SHANNON L. V. and M. J. O'TOOLE (Convenors) 1998 - Proceedings of the International Symposium on Environmental Variability in the South-east Atlantic, Swakopmund, Namibia, 30 March-1 April 1998.
SHANNON, L. V. and S. C. PILLAR 1986 - The Benguela ecosystem. 3. Plankton. In Oceanography and Marine Biology. An Annual Review 24. Barnes, M. (Ed.). Aberdeen; University Press: 65-170.
SHILLINGTON, F. P. 1998 - The Benguela upwelling system off Southwestern Africa, Coastal segment (16, E). In The Sea Vol II, Robinson, A. R. and K. H. Brink (Eds): 583-604.
SILVA, E. S. 1953 - Red water por Exuviaela baltica Lohm. com simultnea mortandade de peixes nas guas litorair de Angola. Ann Junta Invest. Ultramar 8(2).
STANDER, G. H. 1964 - The Benguela Current off South West Africa. Investl. Rep. Mar. Res. Lab. SW Afr. 12: 43 pp plus 77 pages figures.
STANDER, G. H. and A. H. B. DE DECKER 1969 - Some physical and biological aspects of an oceanographic anomaly off South Wets Africa in 1963. Ivestl. Rep. Div. Sea Fish. S. Afr. 81: 46 pp.
STRAMMA, L. and R. G. PETERSON 1989 - Geostrophic transport in the Benguela Current region : J. Phys. Oceanogr. 19: 1440-1448.
TAUNTON-CLARK, J. and L. V. SHANNON 1988 - Annual and interannual variability in the South-east Atlantic during the 20th century. S. Afr. J. mar. Sci. 6: 97-106.
TYSON, P. D. 1986 - Climate Change and Variability in Southern Africa, Oxford univ. press, Cape Town: 220 pp.
VAN BALLEGOOYEN, R. C., GRUNDLINGH M. L. and J. R. E. LUTJEHARMS 1994 - Eddy fluxes of heat and salt from the south west Indian Ocean into the south east Atlantic Ocean : a case study. J. Geophys. Res. 99 (C7) : 14053-14070.
VAN DER LINGEN, C. D. 1994 - Effect of particle size and concentration on the feeding behaviour of adult pilchard, Sardinops sagax. Mar. Ecol. Prog. Ser. 109: 1-13.
VAN DER LINGEN, C. D. 1995 - Respiration rate of adult pilchard Sardinops sagax in relation to temperature, voluntary swimming speed and feeding behaviour. Mar. Ecol. Prog. Ser, 129: 41-54.
VERHEYE, H. M., RICHARDSON, A. J., HUTCHINGS, L., MARSKA, G. and D. GIANAKOURAS 1998 - Long-term trends in the abundance and community structure of coastal zooplankton in the southern Benguela system, 1951-1996. S. Afr. J. mar. Sci. 19 (in press).
VOITUREIZ, B. and A. HERBLAND 1982 - Comparison des systemes productifs de l'Atlantique Tropical Est.: dômes thermiques, upwellings côtiers et upwelling équatorial. Rapp P.V. Reun Con. int. Explor. Mer. 180: 114-130.
WALDRON, H. N. and T. A. PROBYN 1992 - Nitrate supply and potential production in the Benguela upwelling system. In Benguela Trophic Functioning, Payne, A. I. L., Brink, K. H., Mann, K. H. and R. Hilborn (Eds). S. Afr. J. mar. Sci. 12 : 865-871.
WALDRON, H. N., PROBYN, T. A. and G. B. BRUNDRIT 1998 - Carbon pathways and export associated with the southern Benguela upwelling system : A re-appraised. S. Afr. J. mar. Sci. 19 (in press)
WATTENBERG, H. 1938 - Die Verteilung des Sauerstoffs im Atlantischen Ozean. Wiss. Ergeb. Dt. Atlant. Exped. "Meteor" 1925-1927 9: 132 pp.
APPENDICES
APPENDIX I
INSTITUTIONAL INFRASTRUCTURE AND CAPACITY
Human capacity
Hempel and Haslund (1997) have provided a comprehensive assessment of the state of marine science and science capacity in Angola. The Instituto de Investigacao Pesqueira (IIP) is the central institute for basic marine environmental and fisheries research. With a total staff of about 220, of which approximately 40 are university trained, IIP's primary focus is on the monitoring, research and assessment of the fish resources per se. The environmental (oceanography) component of IIP comprises 14 staff members, engaged in various aspects of fisheries oceanography, environmental monitoring and pollution control. The policy of IIP is to increase the skill level of staff, and considerable success has been achieved through close collaboration with the Agostinho Neto University in Luanda and other SADC universities (e.g. University of Cape Town) as well as with universities in Europe, principally in Russia, Germany and Norway. Participation in IOC- sponsored and BENEFIT training initiatives is likewise proving to be beneficial. In spite of difficult operating conditions, with severe limitations in terms of equipment and infrastructure, IIP staff do not lack initiative and enthusiasm, and this bodes well for the future. Donor support from countries such as Norway, Germany and Sweden to strengthen human capacity has been and continues to be vital.
Namibia has a long tradition in marine science and technology, and following independence in 1990, the government has placed a strong emphasis on the training of Namibians and the development of local expertise. The Ministry of Fisheries and Marine Resources (MFMR) is the agency responsible for monitoring, research, assessment, management and control of living marine resources within Namibia's EEZ. The monitoring, research and assessment component, which will be referred to loosely as "research" is headed by a Director based in Windhoek and with most research staff located in Swakopmund at the National Marine Information and Research Centre (NatMIRC). A smaller research group is based in Lüderitz. MFMR has a research and support staff complement of about 100 persons of whom approximately 40 have four-year or higher university qualifications. On paper the "environmental research" component comprises 9 research posts and two technical positions. Recent restructuring within the Ministry in order to strengthen capacity to address pressing fishery monitoring and assessment problems, has de facto resulted in a substantial shrinkage of the oceanography group, although the Ministry is evidently taking steps to redress this. The environmental group, although small, has since independence embarked on a comprehensive monitoring programme, and through close collaboration with, and with strong support from Norwegian and German scientists developed a high degree of competence. (This is reflected in the large number of reports and publications which have been produced by the oceanographers during recent years). Several of MFMR staff have post-graduate qualifications gained at overseas and SADC universities, and like IIP, MFMR has a strong education and training policy. The advent of BENEFIT is enabling further development of Namibian marine science capacity. The Ministry has wisely decided to make maximum use of remotely sensed marine environmental data, and is currently developing appropriate skills to utilise and interpret these. There remains, however, a shortage of technical posts and trained oceanographic technicians. The University of Namibia (UNAM) has a close association with MFMR and is keen to develop collaboration training courses in oceanography and fisheries science. UNAM does not yet offer post-graduate degree courses in marine science.
South African marine science celebrated its centenary in 1995, and can generally be considered as world class. The Marine and Coastal Management (MCM) of the Department of Environmental Affairs and Tourism, based in Cape Town has a total research staff complement of about 150, of whom approximately 50 are university graduates. The support staff comprise highly skilled engineers, technicians and technical assistants. All of the technicians possess formal technical qualifications (NDT oceanography) or similar - equivalent to a three year university degree). The majority of the research staff possess post-graduate degrees. MCM has a long and close association with the University of Cape Town (UCT) and other South African universities, and also since the early 1960s, with the Cape Technikon. Approximately 20 oceanographers and 20 technical staff at MCM are involved in environmental or ecosystem-type research or monitoring, and are divided into "Environment" and "Whole Systems" groups. These provide the disciplinary homes for physical oceanographers, marine chemists, planktologists, ecosystem-expert and modellers. Scientific output as evident in publications in international scientific journals and by way of management advice is high, comparable with leading marine institutes in Europe and North America. Apart from MCM, staff and students of several departments at UCT and the University of the Western Cape (UWC), employees of the South African Museum, CSIR, Institute of Marine Technology
inter alia are engaged in research and monitoring of various aspects of the Benguela environment. At UCT, for example, four full time academic appointees in the Oceanography Department and two in the Zoology Department can be regarded as specialists on the Benguela ecosystem. In addition, the Zoology Department has a long tradition and considerable expertise in inshore ecology. The highly regarded Benguela Ecology Programme served as the principal catalyst for the development of Benguela environmental expertise in South Africa during the 1980s. BENEFIT and BCLME have the potential to do the same for the three countries of the Benguela region over the next few decades.
Infrastructure
(a) Accommodation
The accommodation available for marine science and technology in the region is generally good. In Angola, the headquarters of IIP in Luanda are housed in a large modern building equipped with six laboratories for fisheries/oceanography and food technology, 60 offices for administration staff and scientists, a library and three meeting rooms. Regional offices/laboratories exist in Benguela, Lobito, Namibe and Tombua. (The accommodation at Namibe is new and well designed, and is situated near the harbour. Namibe is likely to be an important centre in Angola for BENEFIT and BCLME activities.) The Agostino Neto University is located in the centre of Luanda. Buildings are in poor condition.
Office and laboratory facilities in Namibia exist at NatMIRC in Swakopmund and in Lüderitz. The former are modern of a high standard, and provide a good working environment for Namibia's marine and fisheries research. Meeting facilities are excellent. Overall the accommodation is very good. The Lüderitz facility is smaller, but has recently been upgraded and is also regarded as very good. The University of Namibia is housed in modern buildings on the outskirts of Windhoek. Office/laboratory accommodation ranges from adequate to excellent.
In South Africa MCM is housed in a commercial building close to the Cape Town city centre. Although not ideal in layout (design) the offices and laboratories are of a relatively high standard and provide a generally satisfactory working environment for MCM staff. Meeting facilities are good and there is a small multi-purpose auditorium. The University of Cape Town is situated 4km away at Rondebosch. Offices and laboratoriums in the Zoology and Oceanography Departments are good. Lecture and seminar rooms provide good venues for meetings and conferences. Excellent library facilities exist at MCM and UCT. The CSIR is situated at Stellenbosch, some 50km from Cape Town. Office and laboratory accommodation there is excellent. The University of the Western Cape is at Bellville, about 20km from Cape Town. Office and laboratory accommodation at UWC is good.
(b) Research Ships
Angola: R. V. Goa. Small, poorly equipped, and not operational
Namibia: R. V. Welwitschia (47m). Modern multi-purpose research stern trawler built in 1994, carries a maximum of 9 scientists, although accommodation for only 5 can be regarded as satisfactory. Laboratories are small. The vessel is not ideally suited to use on multi-disciplinary fish-environment surveys. R. V. Kuiseb (19m). General purpose wooden vessel built in 1958, carries 4 scientists. A very basic replacement is under construction.
South Africa: F. R. S. Africana (78m). Modern all weather multi-purpose fisheries and oceanography research stern trawler built in 1981. Excellent laboratories. Carries 17 scientists. F. R. V. Algoa (53m). Converted commercial stern trawler, with similar capabilities, but on a lesser scale, to Africana . Carries 12 scientists. F. R. V. Sardinops (37m). Old (1958) research side trawler. Limited capability. Carries 4 scientists. The manning arrangements for South African research vessels are problematic.
Norway: R. V. Dr Fridtjof Nansen (78m). Modern multi-purpose fisheries stern trawler. Excellent. Carries 17 scientists. The vessel built in 1992 replaced the previous one of same name which had been operating in the South-east Atlantic since the late 1980s. This is probably the most versatile and useful vessel in the region and the cornerstone of fisheries research (surveys) off Namibia and Angola.
Germany: Although there is no German research ship dedicated to the region, Germany chartered a vessel in 1997 for the investigation of the Angola/Benguela front, and plans to charter vessels for work in the Benguela ecosystem again in the future.
(c) Equipment/Instrumentation
Except R. V. Goa, R. V. Kuiseb and F. R. V. Sardinops, the research ships are well equipped and possess modern instrumentation and gear. In the case of South Africa much of the instrumentation has been locally developed (a result of sanctions) and has during the past five years become run-down as a consequence of shrinking budgets and increased operating costs. Shore-based laboratory facilities in terms of instrumentation and equipment in South Africa and Namibia range from adequate to excellent. The Angolan laboratories are generally poorly equipped, with very limited instrumentation. The supporting infrastructure is likewise totally inadequate. A satellite communication link with IIP in Luanda has recently been established.
APPENDIX II
OCEANOGRAPHIC AND FISHERIES DATA AND INFORMATION SYSTEMS
Angola
There are four separate fisheries data processing systems in operation in Angola: one system is based at the Fisheries Research Institute (IIP), one is in the Directorate of the National Fisheries (DNP), one is at the Directorate of Surveillance (DNF), and there is a system used by the Planning Office in the Ministry of Fisheries (GEP).
The DNP system has been developed through Swedish support and is based on the Swedish Baltic Sea fisheries data system. It is used for commercial fisheries statistics. The version in use in the DNP has been modified to meet local needs, but is as yet incomplete. The system enables the registration of fishing companies, catch discharges, gear and species data, registration of gear in use in the fishery. These data are available for both national, joint venture and foreign fishing vessels and companies. The system also facilitates the registration of fishing enterprises, taxes, contracts, export data and transhipment details. The analysis output is limited at present but the intention is to access and output details on quotas and TACs. Exchange rate updating will be included so that values can reflect local currency fluctuations. The DNP has three MS-Dos Windows based PCs which are as yet unlinked. A small dedicated computer network has recently been purchased. created to deal specifically with the database, with its own personnel and resources.
Angola has recognised that there needs to be a unified data system and has proposed modifying the DNP system so that more detailed analysis for stock and catch assessment can be achieved. A programme to unify the systems has been devised, and this will be done in conjunction with an upgrading of the data collection system. IIP is developing its own database for fisheries and oceanographic scientific data. A new section in IIP has been created to deal specifically with the database, with its own personnel and resources. The oceanographic data still need to be incorporated in the database. A major task will be to recover and include the historic records which span nearly 30 years.
Apart from oceanographic data collected by Angola e. g. a 30 year time series of sea surface temperature, salinity and oxygen at Lobito and other pre- and post-independence records, IIP has a comprehensive set of data from the monitoring and research programme. IIP also holds Russian data sets on the oceanography of the Angola/Benguela front and Angola Dome, while more data sets rest in Kaliningrad.
Namibia
The Ministry of Fisheries and Marine Resources has developed and implemented an integrated Fisheries Information Management System that provides a data capture, information management and reporting system for all core information on the fisheries sector. The FIMS provides for the following modules; Species, Vessels, Factories, Fisheries, Exploitation Rights, Quotas, Payments, Landings, Logsheets and a Management Information System (MIS). This core system is designed to manage all information from and to the fishing industry. Work is also well advanced in the development of two further sub-systems: the Biological, Oceanographic and Supporting Research Module (BOSRM) and the Surveillance and Enforcement Module (SEM). The overall system will thus provide access to the full range of information available tot the Ministry, including all available historical data and international databases. The BOSRM will provide access to, and be integrated with, fisheries sector information to enable accurate and meaningful analysis and interpretation of catch information against fishery and environmental dynamics. However, delays in the implementation of BOSRM mean that there is no viable oceanographic data base available to MFMR scientists, and data are currently stored on disk and hard copy.
It is planned that all information, subject to confidentiality and security, will be made available for research and general use by government, industry, international collaborating partners and agencies through a variety of systems of access.
The system was developed using S-Designer and Powerbuilder and is implemented using Microsoft NT and SQL Server in a Windows 3.1 and 95 environment. There are four sites each with their own server networks at Windhoek (Ministry of Fisheries and Marine Resources HQ), Swakopmund (National Marine Information and Research Center), Walvis Bay (Inspectorate and Operations Centre) and Lüderitz (Inspectorate and Research). These four LANs are connected by a WAN through a leased line system on fibre-optic cables and all network management is from the Information Systems Division in Windhoek. The majority of hardware has been specifically purchased for the designed task (Olivetti servers and work stations) but with a view to appropriate expansion - particularly of storage and processing power.
Namibian oceanographic data holdings, pre-and post-independence, including extensive data collected during cruises of the Dr Fridtjof Nansen during the 1980s and 1990s and during cruises of R. V. Welwitschia and R. V. Matsuyama Maru (Japanese Overseas Fisheries Cooperation Foundation), are extensive and generally of a high quality. Cruise data are augmented by data from coastal monitoring stations (Swakopmund and Lüderitz) and satellite imagery.
South Africa
(a) MCM has been using the Data General range of mini computer since 1979 for the processing of data relating to commercial catches, research samples and environmental parameters. The applications were developed in Cobol and the data stored in an Infos hierarchical database. These systems are currently being converted in order to make use of the advanced features offered by new hardware and software technology. These applications will be run on a local area network using Novel NetWare. The data are being converted to a relational database which will support the integration of common entities, such as grids, vessels and species codes, and provide better facilities for ad hoc management and research queries. The software is being developed using Borland Delphi, a fourth generation language which runs on PCs using Windows 95. The bulk of the systems relate to commercial catches from the following sectors: Demersal, Pelagic, Rock Lobster, (West Coast, South Coast and Natal), Abalone, Linefish and Netfish. Some data sets cover catches from 1978 to date. The environmental parameters (physical and chemical0 and stored in the Oceanographic Database. The issuing of vessel licences and fishing permits is handled by the Boat Registration System.
(b) The University of Cape Town is well connected in the academic and research environments in Southern Africa. Not only does the University have a wide range of computers available, from Macs to PCs to Workstations, but it has expertise that is of world class quality. All of the University computers are connected to the Internet as well as on several local area networks on campus. The University is constantly seeking to upgrade and improve the quality of this interconnectivity so as to increase the flow of information. Through the Centre of Marine Studies and the various Departments at the University, expertise is readily available in the fields of data processing and statistics, resource assessment, management and conservation as well as environmental evaluations and oceanographic remote sensing. These skills can be applied to the problems of data and information management as well as in the interpretation, presentation and publication of the necessary data.
(c) South Africa possesses an extremely comprehensive collection of oceanographic data; discrete, profile and time series. Most of these are available from SADCO (see below) and/or MCM. South Africa also has extensive holdings of NOAA satellite sea surface temperature data.
South African Data Centre for Oceanography (SADCO)
SADCO was established as a national oceanographic data bank in the 1970s to service South Africa's marine science community. It has subsequently developed into a regional facility. SADCO archives, extracts and manipulates oceanographic data from the southern African marine environment and provides a spectrum of professional cost-efficient and user-friendly services. It also promotes the scientific and commercial application of oceanographic data.
SADCO receives data for the area 0°-70°S and 30°W-70°E from a variety of sources including various southern African marine agencies, and World Data Center, and other international data sources by exchange or purchase.
The SADCO data base contains observations since 1850 which include inter alia the following:
-
Oceanographic station data for surface and serial depths, giving values of temperature, salinity, sound velocity, oxygen, nutrients etc.
-
Digital bathythermograph and XBT data
-
Surface data from voluntary observing ships (VOS) including waves, wind and weather, comprising some 3 million readings
SADCO also has access to various document information systems which enable literature searches for published oceanographic data
SADCO is guided by a Steering Committee and managed by CSIR on behalf of its sponsors which include the South African Department of Environmental Affairs and Tourism, the South African Navy, CSIR, Foundation for research Development and the Namibian Ministry of Fisheries and Marine resources. Service charges are modest. SADCO could play an important role in BCLME.
Data Confidentiality/Restrictions
Most oceanographic data collected by regional scientists/research institutions is generally available to other bona fide scientists subject to certain conditions. These conditions include
inter alia appropriate acknowledgement of data ownership, a time clause to give the owner a reasonable period to analyse the data and publish research results. Oceanographic collected in international waters around southern Africa by overseas oceanographic institutes is likewise generally readily available to scientists and technicians in southern African states. Again the principals and ethics of international science and/or intellectual property rights apply. For example, SADCO receives and archives various forms of oceanographic data but can put a limited-period hold on data if requested by the collector of the data to do so. These data would then not be released to a third party during the embargo period unless authorised by the data owner, but the data could be used with other data in the data-base for averaging purposes. The present system works well.
Resource-based data are, however, in a different category and fisheries data may not be readily available outside of the organisation responsible for its collection. This practice is not peculiar to southern Africa and applies almost universally. There are two main reasons for this viz. (a) commercially sourced fisheries data contains information which may give the supplier (fisherman or fishing company) a competitive edge over rivals and (b) raw fisheries data may be regarded as strategic information by management agencies and national governments. Fisheries data are, however, generally readily available in processed form. Nevertheless the fact remains that most fisheries and oceanographic data are collected by, or at the behest of, organisations funded by national tax payers and there is a universal movement towards increased transparency and accountability of governments.
APPENDIX III
LIST OF PERSONS CONSULTED
The major part of this Overview was prepared at the University of Cape Town. It draws on input from a large number of individuals, by way of written documents (reports, publications), or from presentations at symposia and workshops (e. g. International Symposium and Workshop on Environmental Variability in the South-east Atlantic, Swakopmund, March/April 1998; First Regional Workshop on BCLME, Cape Town, July,1998) and from discussions with key role players. The persons consulted specifically for the Overview were as follows:
Dr J. Augustyn (Acting Director, MCM)
Dr L. Hutchings (Chief Specialist Scientist, MCM and Member: BCLME Management Committee)
Dr M. J. O'Toole (MFMR, and BCLME Project Coordinator)
Mr G. W. Bailey (MCM)
Dr R. G. Barlow (MCM)
Dr A. Cockcroft (MCM)
Dr R. J. M. Crawford (MCM)
Dr J. David (MCM)
Prof. J. Field (Zoology Department, UCT)
Dr V. Filipe (IIP)
Mrs P. Krohn, (Oceanography Department, UCT)
Prof. J. R. E. Lutjeharms (Oceanography Department, UCT)
Dr V. de Barros Neto (IIP)
Dr A. Pereiro (IIP/BENEFIT)
Dr S. Pillar (MCM)
Dr G. Pitcher (MCM)
Dr M. de Lourdes Sardinha (IIP)
Ms L. J. Shannon (MCM)
Prof. F. Shillington (Oceanography Department, UCT)
Mr A. P. van Dalsen (MCM)
Dr H. Verheye (MCM)
Dr H. Waldron (Oceanography Department, UCT)
Mr B. Wessels (MCM)
Drs P. Freon, P. Curie and C. Roy (ORSTOM, seconded by the French Government to work in the Benguela region 1998-2000)
In addition to the above, discussions were held with key staff of MFMR in Namibia as part of a separate contract in 1997/1998 funded by The World Bank with the purpose of developing an environmental strategy for MFMR. The principal individuals consulted during this exercise were:
Dr B. Oelofsen (Director: Resource Management MFMR)
Dr. B. van Zyl (Deputy Director: Applied Research MFMR)
Ms J. Botha (MFMR)
Mr C. Bartholomae (MFMR)
Ms A. Risser (MFMR)
Ms K. Noli-Peard (MFMR)
Mrs B. Currie (MFMR)
Mr C. Beyers (MFMR)
Mr A. Kemp (MFMR)
Dr G. Ohe (GTZ)
Dr M. F. Tejedor (seconded to MFMR by Spanish Government)
APPENDIX IV
ACRONYMS
AABW Antarctic Bottom Water
AAIW Antarctic Intermediate Water
BCLME Benguela Current Large Marine Ecosystem
BENEFIT Benguela-Environment-Fisheries-Interaction-Training (Programme)
BEP Benguela Ecology Programme
CZCS Coastal Zone Colour Scanner
EEZ Exclusive Economic Zone
ENSO El Nińo - Southern Oscillation
EU European Union
FRD Foundation for Research Development (South Africa)
GDP Gross domestic Product
GEF Global Environmental Facility (World Bank)
GLOBEC Global Ocean Ecosystem Dynamics
GTZ Deutsche Gesellschaft fur Technische Zusammenarbeit
ICEIDA Icelandic International Development Agency
IGBP International Geosphere-Biosphere Programme
IHDP International Human Dimensions Programme
IIP Instituto de Investigaçao Pesqueira (Angola)
IOC Intergovernmental Oceanographic Commission
MCM Marine and Coastal Management (South Africa)
MFMR Ministry of Fisheries and Marine Resources (Namibia)
NatMIRC National Marine Information and Research Centre (Namibia)
NORAD Norwegian Agency for Development Cooperation
PDF Programme Development Fund (of GEF)
RV/S Research Vessel/Ship
SADC Southern African Development Community
SADCO South African Data Centre for Oceanography
SANCOR South African Network for Coastal and Oceanic Research
SAP Strategic Action Plan
SeaWiFS Sea-viewing Wide Field-of-View Sensor
SFRI Sea Fisheries Research Institute (South Africa)
SIDA Swedish International Development Agency
SPACC Small Pelagic Fish and Climate Change Programme (of GLOBEC)
SST Sea Surface Temperature
STSW Subtropical Surface Water
TAC Total Allowable Catch
TW Thermocline Water
UNAM University of Namibia
UCT University of Cape Town
UNESCO United Nations Education, Scientific and Cultural Organisation
UWC University of the Western Cape
WCRP World Climate Research Programme
APPENDIX V
NOTES ON ANGOLAN OCEANOGRAPHIC CRUISE AND DATA REPORTS AND PUBLICATIONS
Mr A. F. Pereira, a member of the scientific staff of IIP who is currently on secondment to the BENEFIT Secretariat, was commissioned to compile information about oceanographic cruises undertaken in Angola's EEZ and to extract pertinent information from the various cruise and data reports and publications. In collaboration with IIP staff, in particular Mr V. L. L. Filipe, Mr Pereira produced a comprehensive draft document which contains a wealth of information relevant to BCLME, and which will be extremely useful for planning purposes. The following is a short comment on the available information and its potential utility.
Since 1985, the R. V. Dr Fridtjof Nansen has undertaken numerous surveys of Angola's fish resources in collaboration with IIP. During these cruises environmental measurements (e.g. temperature, salinity, dissolved oxygen, plankton, currents etc) were made. Some 18 cruise reports spanning the period 1985-1998 are now available, and these contain information about the various parameters measured, and useful comments on the state of the environment by the compiler, key extracts of which are contained in Mr Pereira's report ("pertinent comment by compiler"). Apart from the R. V. Dr Fridtjof Nansen, cruises were also undertaken by Angola's R. V. Goa and various other research vessels - Russian, Portuguese, Cuban etc. Eleven cruise reports emanating from the R. V. Goa and nine from other vessels are available.
Since Independence, IIP scientists have authored 17 environmental scientific papers. These are mostly in Portuguese, but a number of recent articles are in English. Although it has not been possible in view of time limitations to undertake a comprehensive assessment of these publications, it is clear from the information provided by Mr Pereira that as a set they do provide a valuable addition to the literature on the Benguela ecosystem. What is also apparent is that IIP scientists, drawing on this material and their knowledge of the Angolan marine environment, will be able to make a substantial contribution to the execution of the BCLME during the implementation phase.
FIGURE CAPTIONS
Fig. 1 Locator map and principal bathymetric features of the Benguela region
Fig. 2 Monthly windstress along the west coast of southern Africa, 75km offshore (modified from Boyd 1987). Note the windy areas near Lüderitz and Cape Frio and the pronounced seasonality in the south
Fig. 3 Conceptual picture of wind-induced coastal upwelling (modified from Shannon 1989)
Fig. 4 Satellite-derived sea surface temperature distribution in the southern Benguela, 9 October 1997
Fig. 5 Characteristic temperature-salinity relationships for the South-east Atlantic Ocean and Benguela region
Fig. 6 Vertical east-west sections at three locations in the Benguela, illustrating the depths of principal water masses, viz Tropical Surface Water (TSW), Subtropical Surface Water (STSW), Thermocline Water (TW), Antarctic Intermediate Water (AAIW), North Atlantic Deep Water (NADW) and Antarctic Bottom Water (AABW). Note the absence of AABW in the area north of the Walvis Ridge, and the strongly stratified surface layer off Angola
.
Fig. 7 Physical boundaries of the Benguela and surface (upper layer) currents
Fig. 8 Satellite-derived sea surface temperature distribution in the northern Benguela,
10 April 1997
Fig. 9 Conceptual diagram showing location of zones of formation of oxygen poor water in the South-east Atlantic
Fig.10 A two-dimensional network of nitrate (and hence carbon) pathways between ocean, shelf and sediments. Numbers are grams carbon x 1013 . (Courtesy Dr H. Waldron, Oceanography Department, University of Cape Town)
Fig. 11 Ocean colour image showing chlorophyll distribution in the Benguela obtained
by SeaWiFS satellite, 9 March,1998 - Courtesy NASA and Ocean Space CC
Fig. 12 Bloom of red tide organism Noctiluca scintilans in the southern Benguela
(Courtesy D. Horstman, SFRI)
Fig. 13 Diagram showing partitioning of planktonic food between sardine and anchovy
(from Van der Lingen 1994)
Fig.14 Southern Benguela upwelling foodweb models (a) Rythers (1969) model and (b)
a revised more appropriate model incorporating the microbial foodweb (from
Moloney 1992)
Fig.15 Distribution of sea surface temperature in the northern Benguela, early in 1984,
showing the southward penetration of warm water during a Benguela Nińo (NOAA image generated by and reproduced courtesy of Ms Scarla Weeks, Oceanography Department, University of Cape Town)
Fig.16 Mass mortality of rocklobsters at Elands Bay in the southern Benguela as a
consequence of the appearance of oxygen deficient water (Courtesy Dr A.
Cockcroft, SFRI)
Fig.17 The "Great Ocean Climate Conveyor Belt" (modified from Broecker, 1991)
|