Appendix 8 workshop 1
Ahmet E. Kideys1, Galina A. Finenko2, Boris E. Anninsky2, Tamara Shiganova3,
Abdolghasem Roohi4, Mojgan Rowshan Tabari4, M. Youseffyan4, Mohammad T. Rostamiya4
1Institute of Marine Sciences, Erdemli, Turkey (kideys@ims.metu.edu.tr)
2Institute of Biology of Southern Seas, Sevastopol, Ukraine (shulman@iuf.net)
3P.P.Shirshov Institute of Oceanology RAS, Moscow, Russia
(shiganov@chip.sio.rssi.tr)
4Mazandaran Fisheries Research Center, Sari, Iran (roohi_ark@yahoo.com)
The alien ctenophore Mnemiopsis, native to the eastern coasts of America had caused great damage and therefore concern in the Black Sea after the late 1980s. This ctenophore was then transported to the Caspian Sea via Volga-Don Canal in late 1990s (Ivanov et al. 2000). The impact of the ctenophore in this new environment already seems very significant: by October 2000, abundance and biomass of fodder zooplankton was reported to decrease 5-6 fold (Shiganova et al. 2001) and by the late summer of 2001, the important pelagic fishery of Iran declined sharply (Kideys et al. 2001). To deal with the problem, Kideys et al (2001) suggested several immediate actions. Among these were laboratory studies with the predator ctenophore Beroe to combat Caspian Mnemiopsis and
performing laboratory experiments with Caspian Mnemiopsis to quantify and evaluate its impact on Caspian ecosystem. .
In order to understand the feasibility of Beroe introduction, as an effective predator on Mnemiopsis, into the Caspian Sea, Beroe has been transported from the Black Sea and Bosporus to the Khazerabad laboratory (Mazandaran) on the Caspian coasts of Iran where several experiments on survival of Beroe and on miscellaneous physiological characteristics (feeding, respiration, reproduction and growth) of both species were performed.
Acclimation of Beroe to low salinity
Beroe generally made up of small individuals (10-40 mm) was transported into Caspian coast of Iran in two batches: For the first batch, thirty Beroe sampled from Sinop, Turkey (southern Black Sea;salinity is about 18 ppt) were transported to the laboratory in Khazerabad (Mazandaran, Iran) in a10 lt jar. The total journey time was about 48 hours for this batch of Beroe. In the second batch about 60 Beroe were collected from Bosporus (salinity around 22 ppt). Journey time was around 24 hours for the second batch.
7 individuals from the first batch were kept individually at 26 oC in conditions of original salinity 18 ppt in 5-1 containers during the first day, next day they were removed to the container with lower salinity water about 16 ppt and than transferred into the container filled totally the Caspian water (12-13 ppt).
A total of 42 individuals sampled from Bosporus (salinity approx. 22 ppt) was acclimatised by lowering salinity gradually in 4 days. During the acclimation, animals fed actively with Mnemiopsis. Our main observations were recorded on the video.
Feeding, respiration and growth rates of Beroe on Caspian Mnemiopsis
A series of feeding experiments was undertaken to measure not only the feeding rate of Beroe on Mnemiopsis but also its digestion time, digestion rate, size preference. Additional experiments were performed in order to clarify whether Beroe would eat other zooplankton.
Under conditions of possible selection (4 different size groups of M. leidyi were offered to B. ovata in the same number simultaneously) Beroe mainly preferred the prey of mean size. Small sized Mnemiopsis comprised insignificant part of daily ration.In the absence of selection (these groups were offered separately in the same biomass) Beroe consumed all sized Mnemiopsis with the same rate. This is important as the main part of Mnemiopsis population in the Caspian Sea is comprised by small ctenophores.
The relationship between specific daily ration (SDR,g/g/day) and Beroe wet weight at 2 food concentrations (I- 1.6; II- 1.0 g/l) could be described with 2 power functions (Fig.1):
I – SDR= 3.184 W-0.841 r2 =0.852
II- SDR = 0.842 W-0.904 r2 = 0.701
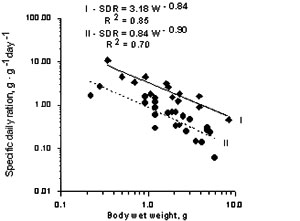
Fig.1. Effect of body weight on specific ration in Beroe ovata at 1.66 g/l (I) and 1.00 g/l (II) food concentration.
The difference between specific daily rations at tested food concentrations showed that food conditions is an important factor in Beroe feeding.
In long-term experiments mean ration of Beroe was 26-43% per body weight on Caspian Mnemiopsis.
Digestion time of B. ovata feeding on M. leidyi at 21± 10C varied from 30 to 450 min in the studied weight range of both ctenophores (13-38 mm in Beroe and 3-27 mm in Mnemiopsis). Interval between two following engulfs usually ranged from 95 to 720 min. Every size of Beroe consumed both small and large M. leidyi; but the ratio between prey and predator weight in these experiments had an insignificant effect on digestion time (Fig.2.) Beroe did not eat Artemia salina or Acartia clausi offered in abundance.
The relationship between oxygen consumption rate (Q, ml O2 /ind/h) and wet weight of B. ovata (g) at 21-230 C is expressed by the equation (Fig.3):
Q= 0.0052 W1.02 r2 = 0.87
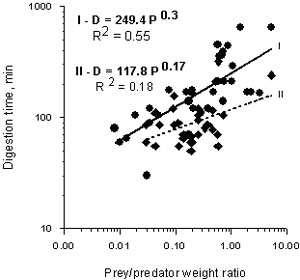
Fig. 2. Effect of prey/predator weight ratio (P) on digestion time (D) in Beroe ovata in Caspian water at 210C (I) and 260C (II).
Value of 1.02 indicates that the weight – specific respiration rate is independent of weight over the measured weight range (0.23-3.87 g).
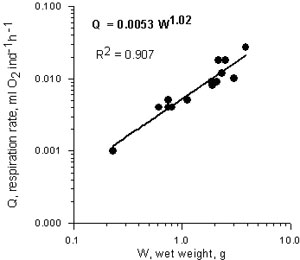
Fig.3. Relationship between respiration rate (ml O2 ind-1h-1) and wet weight (g) in Beroe ovata in the Caspian Sea water.
Analyses of Beroe respiration rate in the Caspian Sea water has shown that Beroe respiration rate is lower of that in the Black Sea (Finenko et al., 2001).
In long-term experiment (12-14 days) average weight for 3 Beroe of the similar initial weight (3.17 g or 30 mm length) increased during the experiment and growth could be expressed with exponential function ( Fig. 4):
W= 2.615 e0.101 t r2 = 0.964, where W is wet weight, g, t- time, days.
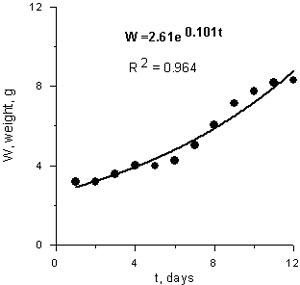
Fig.4. Weight growth of the Beroe ovata in the Caspian Sea water.
The daily specific growth rate of animals according to this equation makes up 0.1 (10%) of body weight. This high value was obtained in conditions of plenty of food and demonstrates high growth potential of these ctenophores. Exponential growth of animals supposes the same specific growth rate in animals of different size (it does not depend on ctenophore weight ); so the population of Beroe in favorable food conditions could be doubled its biomass during 10 days.
The energy budget of B. ovata was calculated (Table 1 ).
Table 1. Daily energy budget (cal/ind/day) of Beroe ovata in the long- term experiment.
|
Initial weight,g |
C |
R |
G |
A |
a |
K1 |
K2 |
|
3.17 |
18.06 |
4.73 |
11.27 |
16 |
0.88 |
0.62 |
0.7 |
|
3.17 |
10.73 |
2.89 |
4.1 |
6.99 |
0.65 |
0.38 |
0.59 |
|
3.17 |
17.11 |
3.57 |
7.7 |
11.27 |
0.65 |
0.45 |
0.68 |
|
Average |
0.72±0.13 |
0.48±0.12 |
0.66±0.06 |
Where C is daily ration, R is respiration rate, G- growth, A – assimilated food in cal/ind/day; a is assimilation efficiency, K1 and K2 – coefficients. Energy content in .M. leidyi is 6.8 cal/g wet weight, B. ovata is 17 cal/ g wet weight.
Mean assimilation efficiency in animals with initial weight 3.17 g was found to be rather high (0.72±0.1) as well as average K1 coefficient (relation between growth and respiration rate) (0.48±0.12) and K2 (relation between growth and assimilated rate) (0.66±0.06). All these values show the high use of consumption and assimilation food for ctenophore growth in conditions of food abundant.
We were able to reproduce Beroe in the laboratory. Thus during experiments 128 eggs of Beroe were obtained and 7 of these were hatched into larvae.
So based on physiological data we could suggest that in the Caspian Sea Beroe is able to ingest Mnemiopsis intensively and decrease its abundance sharply as it happened in the Black Sea
Although we were able to reproduce Beroe into eggs and on a few occasions into larvae, larval development experiments were not successful in the laboratory. However it is still possible that successful development may take place in field conditions
.
Experiments with Mnemiopsis leidyi.
We also quantified different physiological characteristics (feeding, repiration rates and fecundity) of Mnemiopsis leidyi
Clearance rate (volume that Mnemiopsis have to sweep to consume some number of preys) for different weight of Mnemiopsis at 21 0 C increased with increasing of wet weight (Fig.5). The relationship between clearance rate (Cl, ml/ind /h) and wet weight (g) was as following:
Cl= 161.4W0.565 r2= 0.641
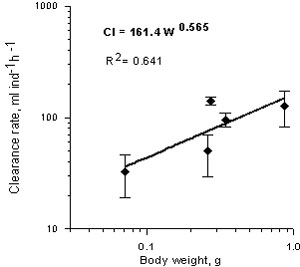
Fig.5. Relationship between clearance rate and body wet weight in Mnemiopsis leidyi in the Caspian Sea.
This clearance rate is higher by a factor of about 2 as compared with this in the Black Sea Mnemiopsis (Finenko & Romanova, 2000).
The relationship between oxygen consumption rate (Q, ml O2 /ind/h and wet weight of M. leidyi (g) at 240 C is expressed by the next equation (Fig.6):
Q= 0.0042W0.774 r2= 0.952
In well adapted Mnemiopsis from the Caspian Sea the respiration rate at ambient temperature (240 C) is 1.5 times as high as this in the Black Sea at the same temperature The difference between oxygen consumption of the Caspian and the Black Sea M. leidyi probably is a result of different food concentration (biomass of zooplankton).
Average fecundity of Mnemiopsis in the Caspian Sea was 1174 eggs/day with maximal 2824 eggs/day for specimens 30-39 mm length and about weight 2.0-2.7 g.
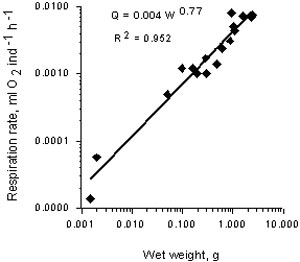
Fig.6. Respiration rate in Mnemiopsis leidyi at 240C in the Caspian Sea
These data will be evaluated with respect to ongoing monitoring studies to evaluate the impact of Mnemiopsis on the pelagic ecosystem of the Caspian Sea in near future.
This study was made possible by the organizational efforts of Dr Vladymyr Vladymyrov (Caspian Environment Program) under financial support of UNOPS, project RER98G32.
References
Finenko G.A. & Z.A Romanova 2000. Population dynamics and energetics of ctenophore Mnemiopsis leidyi in t Sevastopol Bay. Oceanology, v. 40, ?5: 677-685. Translated from Oceanologiya, v.40, No. 5: 720-728.
Finenko G. A., B.E. Anninsky, Z. A. Romanova, G. I. Abolmasova & A. E. Kideys 2001. Chemical composition, respiration and feeding rates of the new alien ctenophore, Beroe ovata, in the Black Sea. Hydrobiologia, 451:177-186.
Ivanov PI, AM Kamakim, VB Ushivtzev, T. Shiganova, O. Zhukova, N. Aladin, SI Wilson, GR Harbison, HJ Dumont 2000. Invasion of Caspian Sea by the comb jellyfish Mnemiopsis leidyi (Ctenophora). Biological Invasions 2: 255-258.
Kideys A. E. , S. Ghasemi, D. Ghninejad, A. Roohi & S. Bagheri 2001. Strategy for combatting Mnemiopsis in the Caspian waters of Iran. Final report prepared for the Caspian Environment Programme, July 2001, Baku, Azerbaijan.
Shiganova TA, AM Kamakin, OP Zhukova, VB Ushivtzev, AB Dulimov, EI Musaeva 2001. An invader in the Caspian Sea: ctenophore Mnemiopsis and its initial effect on pelagic ecosystem. International Workshop on Invasion of Caspian Sea by Mnemiopsis, Baku, Azerbaijan: 24-26.
Document Actions
