|
|
|
ATTACHMENT 4. Development Mnemiopsis leidyi
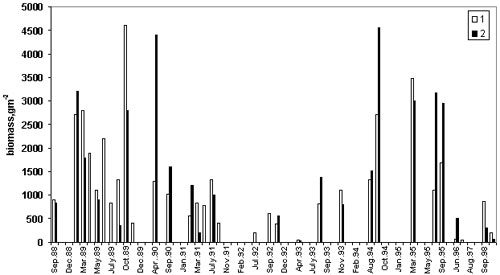
population in the Black Sea and Tamara Shiganova In the early 1980s, M.leidyi was accidentally introduced to the Black Sea. It is a self-fertilizing hermaphrodite, preadapted to rapid colonization in condition of absence of predators it rapidly reached very high abundance. In addition M. leidyi has an ability to regenerate from fragments larger than one quarter of an individual. By summer-autumn, 1988 it was found everywhere, at an average biomass of c. 1 kg WW m-2 (40 g WW m-3) and average abundance of c. 310 m-2 (12.4 m-3) (Vinogradov et al., 1989) (Fig.1). In autumn 1989, the greatest mean biomass ever in the open sea 4.6 kg WW m-2 (184 g m-3 ) and greatest abundance, 7,600 m-2 (304 m-3) were measured In spring 1990, abundance was still very high, but by summer, it began to decrease (Vinogradov et al,1992). This decreasing trend reversed in September 1994, with an average biomass of 2.7 kg WW m-2 (108 g WW m-3) in the open sea, but much higher values in inshore waters (maximum 9.7 kg WW m-2, 176 g WW m-3, average 4.5 kg WW m-2, 180 g WW m-3). A second peak in biomass was observed in spring and summer 1995 (Fig. 1), followed by a second decrease. In 1998, a third increase in offshore waters produced an average biomass of 876 g WW m-2 (35 g WW m-3) and an average abundance of 463 m-2 (18 m-3) (Shiganova, 1998).
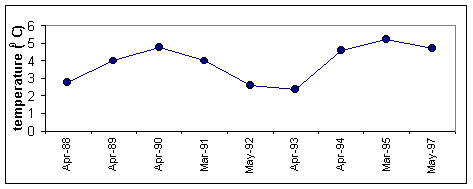
Figure 1. Long-term variation of M.leidyi biomass in the inshore (1) and offshore (2) the Black Sea Vertical distribution M. leidyi mainly lives above the thermocline (from 0 to 15-25 m) during the warm season. Few large specimens venture below the thermocline but remain above the pycnocline (60-80 m). In winter, M. leidyi is found throughout the isothermal layer above the pycnocline, with most of the population above 50 m. Seasonal dynamics and factors controlling population size M. leidyi, as most ctenophores, is annual and does not survive Black Sea winters in areas where water temperatures decrease below 4oC Body size is smaller in spring after a cold winter , and population size after cold winters is low and most often after cold winter follows temperate warm summer and population size of M.leidyi keeps rather low during a year (Fig.2)
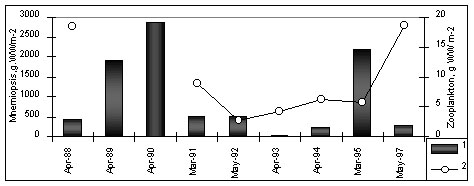
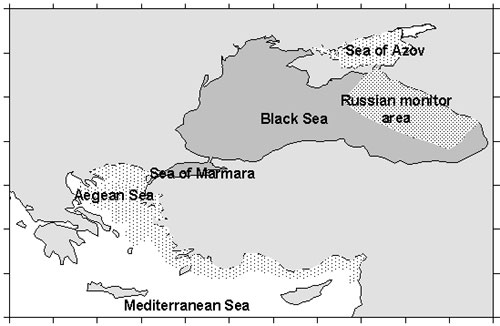
Figure 2. Long-term variation of M.leidyi biomass (1), zooplankton biomass(2) and average temperature air in winter (upper panel) and surface water in August (third panel) Between February and June, overwintering specimens increase by somatic growth only . By June-July, they reach adulthood. Episodic reproduction first begins where zooplankton is abundant and temperature higher than 21oC. Reproduction in inshore waters become intense in mid July to mid August, and continues until October-November. It requires temperatures of 23.5-24oC, and peaks at 24.5-25.5oC. Food supply is also critical , and the main areas of reproduction are inshore waters with abundant Between February and June, overwintering specimens increase by somatic growth only . By June-July, they reach adulthood. Episodic reproduction first begins where zooplankton is abundant and temperature higher than 21oC. Mean individual weight reaches a maximum in July-August, such that biomass peaks in August-September. Density, however, peaks in September-November when reproduction is greatest and the population contains many larvae and small individuals . Reproduction in inshore waters become intense in mid July to mid August, and continues until October-November. It requires temperatures of 23.5-24oC, and peaks at 24.5-25.5oC. Food supply is also critical , and the main areas of reproduction are inshore waters with abundant mesozooplankton in August-September, the time of the so-called “second peak” of zooplankton, composed of warm-water species and larvae of benthic organisms (meroplankton) . However, reproducing ctenophores also spread to the open sea, where spawning continues. During October-November, biomass decreases due to the presence of numerous young individuals (< 3 g WW) and to the death of large individuals after reproduction . In late October to early November, reproduction gradually comes to a standstill, at first in the inshore waters, where temperature drops quickest. Penetration into the other seas of Mediterranean basin M.leidyi from the Black Sea penetrated into the Azov, Marmara, eastern Mediterranean (Fig.3).
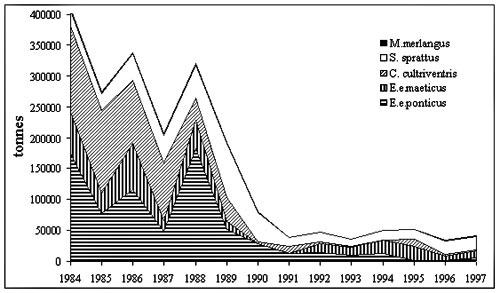
Figure 3. Distribution M.leidyi in the Mediterranean basin The Mediterranean is a system of semi-enclosed seas connected by straits and M.leidyi could penetrated through straits into the adjacent seas. Penetration in the Sea of Azov In 1988 M.leidyi for the first time was found in the Sea of Azov. M.leidyi can survive there only during warm seasons, eliminated in autumn when temperature reachs 40C (Volovik et al, 1993). Penetration in the Sea of Marmara Probably, M. leidyi penetrated the Sea of Marmara with the upper Black Sea current through Bosporus in 1989-1990. It now occurs year-round in the upper water layer. Intense reproduction was recorded in early October 1992, when mean biomass was 4.2 kg.m-2 (152 g WW m-3) and density 27 m-3 (Shiganova, 1993) Distribution in the Aegean Sea and in the eastern Mediterranean Mnemiopsis leidyi was first recorded during late spring-summer 1990 in the Aegean Sea, when it was the most abundant in gulfs and bays ( Saronikos Gulf (45-75 m-2). In subsequent years M.leidyi was found in a few numbers in several coastal areas of the Aegean Sea These specimens M. leidyi may have been carried to the northern Aegean Sea by the Black Sea currents. The size of individuals collected in the Aegean Sea and in the Saronikos Gulf was 2.5–6.5 cm(Shiganova et al, 2001). Further east, M. leidyi appeared in Mersin Bay in spring 1992 (Kideys & Niermann, 1994), and in Syrian coastal waters in October 1993 (Shiganova, 1997). It is even less likely that these specimens were brought with ballast water of the ships as supported by the fact that they were found near Mersin and Latakia ports. Effect on the Black Sea ecosystem The main food of M. leidyi is zooplankton, but fish eggs and larvae (“ichtyoplankton”) directly follow this. The fluctuations between ctenophores and zooplankton were antagonistic (Fig.2). In years with high M. leidyi density, zooplankton biomass had strongly declined by autumn. When M. leidyi density was lower, zooplankton biomass remained higher (Fig. 2). During low ctenophore years, such as 1992-1993 and after 1996, zooplankton, and particularly C. euxinus, showed signs of a recovery. Copepod species diversity increased; some species, which had disappeared in 1990-1992, reappeared. The most striking comeback was that of Paracalanus parvus. There are three predominantly zooplanktivorous fish species in the Black Sea: the Black Sea anchovy (Engraulis encrasicolus ponticus Aleksandrov), Mediterranean horse mackerel (Trachurus mediterraneus ponticus Aleev) and sprat (Sprattus sprattus phalericus Risso, 1827). They became the main commercial species in the 1980s, after the demise of the large pelagic piscivorous fish and dolphins . But their stocks and catches declined dramatically during the bloom of M. leidyi (Fig. 4). The most severe decline was in Black Sea anchovy and Mediterranean horse mackerel, which spawn during summer and suffered from decreased zooplankton abundance. Copepoda had been the main food of Black Sea anchovy, but in 1989 these sharply decreased. Copepods in the diet were replaced for about 30% by larvae of Cirripedia, Ostracoda and Bivalvia, all with a low caloric content.(Fig.5) Consequently, growth rate, weight-at-age, fecundity and frequency of spawning of anchovy decreased (Shiganova & Bulgakova, 2000), while the Mediterranean horse mackerel completely disappeared from Russian commercial catches.
Figure 4. Former USSR catches of zooplanktivorous fish in the Black and Azov Seas
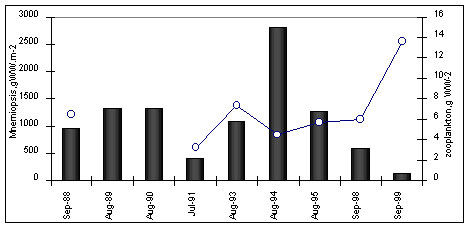
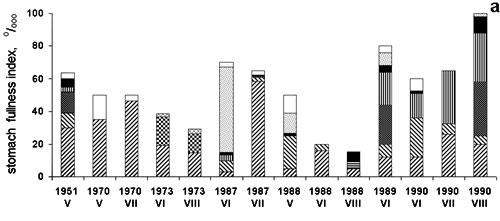
Figure 5. TheBlack Sea anchovy stomach fullness There was also an adverse effect on moderate cold water fish (e.g. sprat), in spite of their spawning in autumn and winter. These co-occurred with M. leidyi in the intermediate layer in winter and only occasionally in summer , but there was effect on the food contents and rations also for sprat.(Shiganova & Bulgakova, 2000). (Fig.6)
Figure 6. Diet composition of sprat M. leidyi affected the food base of all fish larvae. The percentage of starving larvae increased to high values during the bloom of M. leidyi. Due to the absence of small copepod species, larvae had to switch to bigger-sized organisms, which are less suitable and caused a heavy mortality Effect on the the Azov Sea ecosystem was even stronger than in the Black Sea. During the first months of summer M.leidyi consumed almost all of the zooplankton. The stocks of planktivorous fish dropped and recovered little, due to a persistent summer abundance of M. leidyi . The strong effect of M.leidyi on the Black and Azov Seas also reflects an absence of predators. Black Sea mesozooplankton was decimated by M. leidyi. Planktivorous among fish suffered from competition and consumption of their pelagic eggs and larvae enormous numbers of M. leidyi, unchecked by predators of their own. After the appearance of B .ovata, the situation took a new, spectacular turn: M.leidyi plummeted, and the ecosystem began to recover (Shiganova et al. 2000). However during seasonal absence of B.ovata during spring –summer M.leidyi can reach high density and grazes zooplankton to the level before M.leidyi appearance (Fig.7).
Figure 7. Biomass of Mnemiopsis leidyi(1) and Beroe ovata (2) during 1999-2000 in the north-eastern Black Sea Literature Kideys, A. E. & U. Niermann 1994. Occurrence of Mnemiopsis along the Turkish coast. ICES J. mar. Sci.51: 423-427. Shiganova, T. A.,1993. Ctenophore Mnemiopsis leidyi and ichthyoplankton in the Sea of Marmara in October of 1992. Oceanology 33: 900-903. Shiganova, T. A., 1997. Mnemiopsis leidyi abundance in the Black Sea and its impact on the pelagic community. In”Sensivity of North Sea, Baltic Sea and Black Sea to antropogenic and climatic changes” . In E.Ozsoy & A.Mikaelyan (eds), Kluwer Academic Publishers, Dordrecht /Boston/ London: 117-130. Shiganova, T. A, 1998. Invasion of the Black Sea by the ctenophore Mnemiopsis leidyi and recent changes in pelagic community structure. Fish. Oceanogr. 7 –GLOBEC Special Issue Ed.Steeve Coombs:305-310 Shiganova, T. A. & Y. V. Bulgakova, 2000. Effect of gelatinous plankton on the Black and Azov Sea fish and their food resources. ICES J. mar. Sci. 57: 641-648. Shiganova, T. A, J. V. Bulgakova, P. Y. Sorokin & Y. F. Lukashev, 2000. Investigation of a new invader, Beroe ovata in the Black Sea. Biology Bulletin 27: 247-255. Shiganova T. A., Mirzoyan Z. A., Studenikina E. A., Volovik S. P., Siokou-Frangou I., Zervoudaki S., Christou E. D., Skirta A. Y., and Dumont H. J.Development population of the invader ctenophore Mnemiopsis leidyi ( A.Agassiz) in the Black Sea and in other seas of the Mediterranean basin// Marine biology. 2001. In press. Vinogradov, M. E., E. A Shushkina., E. I Musaeva., P. Y. Sorokin,1989. Ctenophore Mnemiopsis leidyi (A.Agassiz) (Ctenophora: Lobata) - new settler in the Black Sea. Oceanology 29:.293-298. Vinogradov, M. E., V. V Sapozhnikov & E. A. Shushkina 1992. The Black Sea ecosystem. Moscow. Russia. Nauka, 112 p. (in Russian). Volovik, S. P., I. A. Mirzoyan & G. S. Volovik, 1993. Mnemiopsis leidyi: biology, population dynamics, impact to the ecosystem and fisheries. ICES. (Biol. Oceanogr. Committee) 69: 1-11.
|
Meeting Report
Attachment
1 Appendix
1
|
|